Perinnöllisen vaihtelun tyypit. Perinnöllinen ja ei-perinnöllinen vaihtelu
Biologian vaihtelu on yksilöllisten erojen syntymistä saman lajin yksilöiden välillä. Vaihtuvuuden vuoksi populaatio muuttuu heterogeeniseksi ja lajilla on paremmat mahdollisuudet sopeutua muuttuviin ympäristöolosuhteisiin.
Biologian kaltaisessa tieteessä perinnöllisyys ja vaihtelevuus kulkevat käsi kädessä. Vaihtuvuutta on kahdenlaisia:
- Ei-perinnöllinen (muunnos, fenotyyppinen).
- Perinnöllinen (mutaatio, genotyyppinen).
Ei-perinnöllinen vaihtelu
Biologian modifikaatiovaihtelu on yksittäisen elävän organismin (fenotyypin) kykyä sopeutua genotyyppinsä sisällä oleviin ympäristötekijöihin. Tämän ominaisuuden ansiosta yksilöt sopeutuvat ilmaston ja muiden olemassaolon olosuhteiden muutoksiin. on missä tahansa organismissa tapahtuvien mukautumisprosessien ytimessä. Joten ulkosiitoseläimissä, kun pito-olosuhteet paranevat, tuottavuus kasvaa: maidontuotanto, munantuotanto jne. Ja vuoristoalueille tuodut eläimet kasvavat lyhyiksi ja niillä on hyvin kehittynyt aluskarva. Myös ympäristötekijöiden muutokset aiheuttavat vaihtelua. Esimerkkejä tästä prosessista löytyy helposti jokapäiväisestä elämästä: ihmisen iho tummuu ultraviolettisäteiden vaikutuksesta, lihakset kehittyvät fyysisen rasituksen seurauksena, varjostetuilla alueilla ja valossa kasvavilla kasveilla on erilaisia lehtien muotoja ja jänikset muuttavat lehtiään. turkin väri talvella ja kesällä.
Seuraavat ominaisuudet ovat ominaisia ei-perinnölliselle vaihtelulle:
- muutosten ryhmäluonne;
- ei ole peritty jälkeläisille;
- muutos piirteen genotyypin sisällä;
- muutosasteen suhde ulkoisen tekijän vaikutuksen voimakkuuteen.
Perinnöllinen vaihtelu
Perinnöllinen tai genotyyppinen variaatio biologiassa on prosessi, jossa organismin genomi muuttuu. Hänen ansiostaan yksilö saa merkkejä, jotka olivat aiemmin epätavallisia hänen lajilleen. Darwinin mukaan genotyyppinen vaihtelu on evoluution päätekijä. Perinnöllistä vaihtelua on seuraavanlaisia:
- mutaatio;
- yhdistelmä.
Se tapahtuu geeninvaihdon seurauksena seksuaalisen lisääntymisen aikana. Samalla vanhempien ominaisuudet yhdistyvät eri tavoin useissa sukupolvissa, mikä lisää populaation organismien monimuotoisuutta. Kombinatiivinen muuttuvuus noudattaa Mendelin periytymissääntöjä.

Esimerkki tällaisesta vaihtelusta on sisäsiitos ja ulkosiitos (läheinen ja ei-liittyvä risteytys). Kun yksittäisen tuottajan ominaisuudet halutaan kiinnittyä eläinrotuun, käytetään läheisesti liittyvää risteytystä. Siten jälkeläisistä tulee yhtenäisempiä ja ne vahvistavat linjan perustajan ominaisuuksia. Sukusiitos johtaa resessiivisten geenien ilmentymiseen ja voi johtaa linjan rappeutumiseen. Jälkeläisten elinkelpoisuuden lisäämiseksi käytetään ulkosiitosta - riippumatonta risteyttämistä. Samalla jälkeläisten heterotsygoottisuus lisääntyy ja populaation monimuotoisuus lisääntyy, minkä seurauksena yksilöiden vastustuskyky ympäristötekijöiden haitallisia vaikutuksia vastaan kasvaa.

Mutaatiot puolestaan jaetaan:
- genominen;
- kromosomaalinen;
- geeni;
- sytoplasminen.
Sukusoluihin vaikuttavat muutokset ovat periytyviä. Mutaatiot voivat siirtyä jälkeläisiin, jos yksilö lisääntyy vegetatiivisesti (kasvit, sienet). Mutaatiot voivat olla hyödyllisiä, neutraaleja tai haitallisia.
Genomiset mutaatiot
Biologian vaihtelu genomisten mutaatioiden kautta voi olla kahden tyyppistä:
- Polyploidia - Mutaatio on yleinen kasveissa. Se johtuu ytimessä olevien kromosomien kokonaismäärän moninkertaisesta kasvusta, joka muodostuu rikkomalla niiden eroa solun napoihin jakautumisen aikana. Polyploidisia hybridejä käytetään laajasti maataloudessa- Kasvinviljelyssä on yli 500 polyploidia (sipuli, tattari, sokerijuurikas, retiisi, minttu, viinirypäleet ja muut).
- Aneuploidia on kromosomien lukumäärän lisääntyminen tai väheneminen yksittäisissä pareissa. Tämän tyyppiselle mutaatiolle on ominaista yksilön alhainen elinkelpoisuus. Ihmisissä laajalle levinnyt mutaatio - yksi 21. parista - aiheuttaa Downin oireyhtymän.
Kromosomimutaatiot
Biologian vaihtelu ilmenee muuttamalla itse kromosomien rakennetta: terminaalialueen menetys, geenisarjan toistuminen, yksittäisen fragmentin kierto, kromosomisegmentin siirtyminen toiseen paikkaan tai toiseen kromosomiin. Tällaiset mutaatiot johtuvat usein säteilystä ja ympäristön kemiallisesta saastumisesta.

Geenimutaatiot
Merkittävä osa tällaisista mutaatioista ei ilmene ulkoisesti, koska se on resessiivinen ominaisuus. Geenimutaatiot johtuvat nukleotidien - yksittäisten geenien - sekvenssin muutoksesta ja johtavat uusien ominaisuuksien omaavien proteiinimolekyylien ilmaantumiseen.

Geenimutaatiot ihmisillä aiheuttavat joidenkin perinnöllisten sairauksien - sirppisoluanemian, hemofilian - ilmenemisen.
Sytoplasmiset mutaatiot
Sytoplasmiset mutaatiot liittyvät DNA-molekyylejä sisältävän solun sytoplasman rakenteiden muutoksiin. Näitä ovat mitokondriot ja plastidit. Tällaiset mutaatiot välittyvät äidin linjan kautta, koska tsygootti vastaanottaa kaiken sytoplasman äidin munasta. Esimerkki biologian vaihtelua aiheuttaneesta sytoplasmisesta mutaatiosta on kasvien höyhenpeitteisyys, joka johtuu kloroplastien muutoksista.
Kaikilla mutaatioilla on seuraavat ominaisuudet:
- Ne ilmestyvät yhtäkkiä.
- Ne ovat perinnöllisiä.
- Heillä ei ole suuntaa. Sekä merkityksetön paikka että elintärkeä merkki voivat muuttua.
- Ne syntyvät yksittäisissä yksilöissä, toisin sanoen ne ovat yksilöllisiä.
- Mutaatiot voivat ilmentyessään olla resessiivisiä tai hallitsevia.
- Sama mutaatio voidaan toistaa.
Jokainen mutaatio johtuu tietystä syystä. Useimmissa tapauksissa sitä ei voida määrittää tarkasti. Koeolosuhteissa mutaatioiden saamiseksi käytetään ulkoisen ympäristön suunnattua tekijää - säteilyaltistusta ja vastaavia.
Ajatella!
Kysymyksiä
1. Mitä kromosomeja kutsutaan sukupuolikromosomeiksi?
2. Mitä ovat autosomit?
3. Mitä on homogameettinen ja heterogameettinen seksi?
4. Milloin geneettinen sukupuolen määrittely tapahtuu ihmisillä ja mistä tämä johtuu?
5. Mitä sukupuolen määrittelymekanismeja tiedät? Antaa esimerkkejä.
6. Selitä, mitä sukupuoleen liittyvä perinnöllisyys on.
7. Miten värisokeus periytyy? Millainen värien havaitseminen on lapsilla, joiden äiti on värisokea ja isällä on normaali näkö?
Selitä geneettisestä näkökulmasta, miksi miehet ovat paljon värisokeampia kuin naiset.
Vaihtuvuus- yksi elävien olentojen tärkeimmistä ominaisuuksista, elävien organismien kyky olla olemassa eri muodoissa, hankkia uusia merkkejä ja ominaisuuksia. Vaihtuvuutta on kahdenlaisia: ei-perinnöllinen(fenotyyppinen tai muunnos) ja perinnöllinen(genotyyppinen).
■ Ei-perinnöllinen (muutos) vaihtelu. Tämäntyyppinen vaihtelu on prosessi, jossa uusia ominaisuuksia ilmaantuu ympäristötekijöiden vaikutuksesta, jotka eivät vaikuta genotyyppiin. Tästä seuraa, että ominaisuuksien muutokset - muunnelmat - eivät periydy. Kaksi identtistä (monotsygoottista) kaksoset, joilla on täysin samat genotyypit, mutta kohtalon tahdosta kasvaneet eri olosuhteissa, voivat olla hyvin erilaisia toisistaan. Klassinen esimerkki ulkoisen ympäristön vaikutuksesta ominaisuuksien kehittymiseen on nuolenpää. Tämä kasvi kehittää kolmenlaisia lehtiä kasvuolosuhteista riippuen - ilmassa, vesipatsaassa tai pinnalla.
Ympäristön lämpötilan vaikutuksesta Himalajan kanin villan väri muuttuu. Kohdussa kehittyvä alkio on korkeissa lämpötiloissa, mikä tuhoaa turkin värjäykseen tarvittavan entsyymin, joten kanit syntyvät täysin valkoisina. Pian syntymän jälkeen tietyt ulkonevat kehon osat (nenä, korvien kärjet ja hännänpäät) alkavat tummua, koska siellä lämpötila on alhaisempi kuin muualla eikä entsyymi tuhoudu. Jos revit pois alueen valkoista villaa ja jäähdytät ihoa, alueelle kasvaa mustaa villaa.
Samanlaisissa ympäristöolosuhteissa geneettisesti läheisissä organismeissa modifikaatiovaihtelulla on ryhmäluonne, esimerkiksi kesällä UV-säteiden vaikutuksesta useimpien ihmisten ihoon kertyy suojaavaa pigmenttiä, melaniinia, ihmiset ottavat aurinkoa.
Yhdessä ja samantyyppisissä organismeissa ympäristöolosuhteiden vaikutuksesta eri ominaisuuksien vaihtelevuus voi olla täysin erilaista. Esimerkiksi iso karjaa maidon tuotto, paino, hedelmällisyys riippuvat erittäin voimakkaasti ruokinta- ja ylläpitoolosuhteista ja esimerkiksi vaikutuksen alaisen maidon rasvapitoisuudesta ulkoiset olosuhteet hyvin vähän muutoksia. Kunkin ominaisuuden modifikaatiovaihteluiden ilmenemismuotoja rajoittaa niiden oma reaktionopeus. Reaktionopeus- nämä ovat rajat, joiden sisällä on mahdollista muuttaa piirrettä tietyssä genotyypissä. Toisin kuin itse modifikaatiovaihtelu, reaktionormi periytyy ja sen rajat ovat erilaiset erilaisia merkkejä ja yksittäisissä yksilöissä. Kapein reaktionopeus on ominaista merkeille, jotka tarjoavat organismin elintärkeitä ominaisuuksia.
Koska suurimmalla osalla modifikaatioista on mukautuva arvo, ne edistävät sopeutumista - organismin sopeutumista normaalilla reaktioalueella olemassaoloon muuttuvissa olosuhteissa.
■ Perinnöllinen (genotyyppinen) vaihtelu... Tämän tyyppinen vaihtelu liittyy genotyypin muutoksiin, ja sen seurauksena hankitut ominaisuudet perivät seuraavat sukupolvet. Genotyyppisen muuntelun muotoja on kaksi: kombinatiivinen ja mutaatio.
Kombinatiivinen vaihtelu koostuu uusien ominaisuuksien ilmaantumisesta muiden vanhempien geenien yhdistelmien muodostumisen seurauksena jälkeläisten genotyypeissä. Tämän tyyppinen vaihtelu perustuu homologisten kromosomien itsenäiseen eroon ensimmäisessä meioottisessa jakautumisessa, sukusolujen sattumanvaraiseen tapaamiseen samassa vanhempaparissa hedelmöityksen aikana ja vanhempien parien satunnaiseen valintaan. Se johtaa myös geneettisen materiaalin rekombinaatioon ja lisää homologisten kromosomien alueiden vaihdon vaihtelua, mikä tapahtuu meioosin ensimmäisessä profaasissa. Siten kombinatiivisen vaihtelun prosessissa geenien ja kromosomien rakenne ei muutu, mutta uudet alleeliyhdistelmät johtavat uusien genotyyppien muodostumiseen ja sen seurauksena uusien fenotyyppien jälkeläisten syntymiseen.
Mutaatiovaihtelu ilmaistuna organismin uusien ominaisuuksien ilmaantumisena mutaatioiden muodostumisen seurauksena. Ensimmäistä kertaa termin "mutaatio" otti käyttöön hollantilainen kasvitieteilijä Hugo de Vries vuonna 1901. Nykyaikaisten käsitteiden mukaan mutaatiot ovat äkillisiä luonnollisia tai keinotekoisesti aiheutettuja muutoksia geneettisessä materiaalissa, jotka johtavat muutokseen organismin tietyissä fenotyyppisissä piirteissä ja ominaisuuksissa. Mutaatiot ovat luonteeltaan suuntaamattomia, eli satunnaisia, ja ne ovat tärkein perinnöllisten muutosten lähde, jota ilman organismien evoluutio on mahdotonta. 1700-luvun lopulla. Amerikassa syntyi lyhytraajoinen lammas, joka synnytti uuden Anconian rodun. Ruotsissa XX vuosisadan alussa. platinanvärinen minkki syntyi turkistilalla. Koirien ja kissojen ominaisuuksien valtava valikoima johtuu mutaatioiden vaihtelevuudesta. Mutaatiot syntyvät harppauksin, kuin uusia laadullisia muutoksia: awnless vehnä muodostui piikavehnästä, lyhyet siivet ja raidalliset silmät ilmestyivät Drosophilassa, kaneissa agoutin luonnollisesta luonnollisesta väristä, mutaatioiden seurauksena valkoinen, ruskea, musta väri ilmestyi.
Alkuperäpaikan mukaan erotetaan somaattiset ja generatiiviset mutaatiot. Somaattiset mutaatiot syntyvät kehon soluissa, eivätkä ne välity sukupuolisen lisääntymisen aikana seuraaville sukupolville. Esimerkkejä tällaisista mutaatioista ovat ikäpilkut ja ihosyylit. Generatiiviset mutaatiot esiintyy sukusoluissa ja periytyy.
Geneettisen materiaalin muutostason mukaan erotetaan geeni-, kromosomi- ja genomimutaatiot. Geenimutaatiot aiheuttaa muutoksia yksittäisissä geeneissä, mikä häiritsee nukleotidien järjestystä DNA-ketjussa, mikä johtaa muuttuneen proteiinin synteesiin.
Kromosomimutaatiot vaikuttavat merkittävään osaan kromosomia, mikä johtaa useiden geenien toiminnan häiriintymiseen kerralla. Erillinen kromosomin fragmentti voi kaksinkertaistua tai kadota, mikä aiheuttaa vakavia häiriöitä kehon toiminnassa aina alkion kuolemaan asti kehityksen alkuvaiheessa.
Genomiset mutaatiot johtaa kromosomien lukumäärän muutokseen meioottisten jakautumien kromosomien eroavaisuuksien rikkomisen seurauksena. Kromosomin puuttuminen tai ylimääräisen läsnäolo johtaa haitallisiin seurauksiin. Tunnetuin esimerkki genomisesta mutaatiosta on Downin oireyhtymä, kehityshäiriö, joka ilmenee, kun ylimääräinen kromosomi 21 ilmaantuu. Tällaisilla ihmisillä kromosomien kokonaismäärä on 47.
Alkueläimissä ja kasveissa havaitaan usein kromosomien lukumäärän kasvua, joka on haploidijoukon monikerta. Tällaista muutosta kromosomijoukossa kutsutaan polyploidia... Polyploidien ilmaantuminen liittyy erityisesti homologisten kromosomien hajoamattomuuteen meioosissa, jonka seurauksena diploidisissa organismeissa ei voi muodostua haploideja vaan diploidisia sukusoluja.
Mutageeniset tekijät... Kyky mutatoitua on yksi geenien ominaisuuksista, joten mutaatioita voi esiintyä kaikissa organismeissa. Jotkut mutaatiot ovat yhteensopimattomia elämän kanssa, ja ne vastaanottanut alkio kuolee kohdussa, toiset aiheuttavat pysyviä muutoksia ominaisuuksissa, jotka ovat vaihtelevissa määrin yksilön elämän kannalta merkittäviä. Normaaliolosuhteissa yksittäisen geenin mutaatiotaajuus on erittäin alhainen (10 -5), mutta on ympäristötekijöitä, jotka nostavat tätä arvoa merkittävästi aiheuttaen peruuttamattomia häiriöitä geenien ja kromosomien rakenteessa. Tekijöitä, joiden vaikutus eläviin organismeihin johtaa mutaatioiden määrän lisääntymiseen, kutsutaan mutageenisiksi tekijöiksi tai mutageeneiksi.
Kaikki mutageeniset tekijät voidaan jakaa kolmeen ryhmään.
Fyysiset mutageenit ovat kaikentyyppisiä ionisoivaa säteilyä (y-säteet, röntgensäteet), ultraviolettisäteily, korkeat ja matalat lämpötilat.
Kemialliset mutageenit- nämä ovat nukleiinihappojen, peroksidien, raskasmetallisuolojen (lyijy, elohopea), typpihapon ja joidenkin muiden aineiden analogeja. Monet näistä yhdisteistä aiheuttavat poikkeavuuksia DNA:n replikaatiossa. Mutageenisia vaikutuksia aiheuttavat maataloudessa tuholaisten ja rikkakasvien torjuntaan käytetyt aineet (torjunta-aineet ja rikkakasvien torjunta-aineet), teollisuusjätteet, tietyt elintarvikevärit ja säilöntäaineet, eräät lääkkeet, tupakansavun komponentit.
Venäjällä ja muissa maailman maissa on perustettu erityisiä laboratorioita ja instituutteja testaamaan kaikkien uusien syntetisoitujen kemiallisten yhdisteiden mutageenisuutta.
4. PERINNÄYTYVÄN MUUTTUVUUDEN ROOLI LAJIEN JA SEN MUOTOJEN EVOLUUTIOESSA
Darwinin evoluutioteoriassa evoluution lähtökohtana on perinnöllinen vaihtelevuus, ja evoluution liikkeellepaneva voima ovat olemassaolon taistelu ja luonnonvalinta. Evoluutioteoriaa luodessaan Charles Darwin viittaa toistuvasti jalostuskäytännön tuloksiin. Hän osoitti, että vaihtelevuus on lajikkeiden ja rotujen monimuotoisuuden taustalla. Vaihtuvuus on prosessi, jossa jälkeläisissä esiintyy eroja esivanhempiin verrattuna, mikä määrää lajikkeen, rodun yksilöiden monimuotoisuuden. Darwin uskoo, että vaihtelevuuden syyt ovat ympäristötekijöiden (suorien ja epäsuorien) vaikutus organismeihin sekä itse organismien luonne (koska jokainen niistä reagoi erityisesti ulkoisen ympäristön vaikutuksiin). Vaihtelevuus toimii perustana uusien ominaisuuksien muodostumiselle organismien rakenteessa ja toiminnoissa, ja perinnöllisyys kiinnittää nämä piirteet.Darwin, analysoiden vaihtelevuuden muotoja, nosti niistä kolme: määrätyn, epämääräisen ja korrelatiivisen.
Tietty tai ryhmävaihtelu on vaihtelua, joka syntyy jonkin ympäristötekijän vaikutuksesta, joka vaikuttaa tasaisesti kaikkiin lajikkeen tai rodun yksilöihin ja muuttuu tiettyyn suuntaan. Esimerkkejä tällaisista vaihteluista ovat painon nousu eläinyksilöiden, joilla on hyvä ravinto, hiusrajan muutos ilmaston vaikutuksesta jne. Tietty vaihtelu on massiivista, kattaa koko sukupolven ja ilmaistaan jokaisessa yksilössä samalla tavalla. Se ei ole perinnöllistä, eli muuttuneen ryhmän jälkeläisillä muissa olosuhteissa vanhempien hankkimat hahmot eivät periydy.
Epävarma eli yksilöllinen vaihtelevuus ilmenee erityisesti jokaisessa yksilössä, ts. sinkku, yksilöllinen luonteeltaan. Se liittyy eroihin saman lajikkeen tai rodun yksilöissä samanlaisissa olosuhteissa. Tämä vaihtelevuuden muoto on määrittelemätön, eli merkki samoissa olosuhteissa voi muuttua eri suuntiin. Esimerkiksi yhdessä kasveissa esiintyy yksilöitä, joilla on eri väriset kukat, erilainen terälehtien värin voimakkuus jne. Syy tähän ilmiöön ei ollut Darwinille tiedossa. Epävarmalla vaihtelulla on perinnöllinen luonne, eli se siirtyy tasaisesti jälkeläisiin. Tämä on sen suuri merkitys evoluution kannalta.
Korrelatiivisella tai suhteellisella vaihtelulla minkä tahansa elimen muutos on syy muiden elinten muutoksiin. Esimerkiksi koirilla, joilla on huonosti kehittynyt turkki, on yleensä alikehittyneet hampaat, kyyhkysillä, joilla on höyhenjalkainen kalvo, varpaiden välissä, kyyhkyillä, joilla on pitkä nokka, on yleensä pitkät jalat, valkoisilla kissoilla siniset silmät yleensä kuuro jne. Korrelatiivisen vaihtelun tekijöistä Darwin tekee tärkeän johtopäätöksen: ihminen, joka valitsee minkä tahansa rakenteen piirteen, melkein "todennäköisesti vahingossa muuttaa ruumiin muita osia salaperäisten korrelaatiolakien perusteella".
Määritettyään vaihtelevuuden muodot Darwin tulee siihen tulokseen, että vain periytyneet muutokset ovat tärkeitä evoluutioprosessille, koska vain ne voivat kertyä sukupolvelta toiselle. Darwinin mukaan päätekijät kulttuuristen muotojen kehityksessä ovat perinnöllinen vaihtelevuus ja ihmisen tekemä valinta (Darwin kutsui sellaista valintaa keinotekoiseksi). Vaihtelevuus on keinotekoisen valinnan välttämätön edellytys, mutta se ei vaikuta uusien rotujen ja lajikkeiden muodostumiseen.
PÄÄTELMÄ
Siten Darwin rakensi ensimmäistä kertaa biologian historiassa evoluutioteorian. Tällä oli suuri metodologinen merkitys, ja se ei vain pystynyt perustelemaan selkeästi ja vakuuttavasti ajatusta orgaanisesta evoluutiosta nykyaikaisille, vaan myös varmistamaan itse evoluutioteorian pätevyyden. Tämä oli ratkaiseva vaihe yhdessä luonnontieteen suurimmista käsitteellisistä vallankumouksista. Tärkeintä tässä vallankumouksessa oli evoluution teologisen ajatuksen korvaaminen alkuperäisen tarkoituksenmukaisuuden käsitteenä luonnonvalinnan mallilla. Kovasta kritiikistä huolimatta Darwinin teoria sai nopeasti hyväksynnän, koska käsitys elävän luonnon historiallisesta kehityksestä oli havaittujen tosiseikkojen selityksessä parempi kuin käsitys lajien muuttumattomuudesta. Teoriansa tueksi Darwin käytti, toisin kuin edeltäjänsä, valtavan määrän tietoja, joita hänellä oli saatavilla eri aloilta. Bioottisten suhteiden ja niiden populaatio-evolutionaarisen tulkinnan esille tuominen oli darwinilaisen evoluutiokäsityksen tärkein innovaatio ja antaa oikeuden päätellä, että Darwin loi olemassaolotaistelun käsityksensä, joka poikkesi olennaisesti edeltäjiensä ideoista. Oppi orgaanisen maailman evoluutiosta oli ensimmäinen kehitysteoria, jonka loi "luonnonhistoriallisen materialismin avulla luonnontieteen syvyyksissä, ensimmäinen kehitysperiaatteen soveltaminen itsenäiseen luonnontieteiden alaan". Tämä on darwinismin yleinen tieteellinen merkitys.
Darwinin ansio ja se, että hän avasi liikkeellepaneva voima orgaaninen evoluutio. Biologian jatkokehitys syvensi ja täydensi hänen ajatuksiaan, jotka toimivat modernin darwinismin perustana. Kaikilla biologisilla tieteenaloilla johtava paikka on nyt historiallisella tutkimusmenetelmällä, jonka avulla on mahdollista tutkia organismien tiettyjä evoluution polkuja ja tunkeutua syvälle biologisten ilmiöiden olemukseen. Charles Darwinin evoluutioteoria on löytänyt laajan sovelluksen modernissa synteettisessä teoriassa, jossa luonnonvalinta on edelleen ainoa ohjaava tekijä evoluutiota, jonka materiaalina on mutaatio. Darwinin teorian historiallinen analyysi synnyttää väistämättä uusia tieteen metodologisia ongelmia, joista voi tulla erityisen tutkimuksen aihe. Näiden ongelmien ratkaiseminen edellyttää tietokentän laajentamista ja siten tieteellistä edistystä monilla aloilla: sekä biologiassa, lääketieteessä että psykologiassa, joihin Charles Darwinin evoluutioteorialla oli yhtä paljon vaikutusta kuin luonnontieteisiin. .
Luettelo käytetystä kirjallisuudesta
1. Alekseev V.A. Darwinismin perusteet (historiallinen ja teoreettinen johdanto). - M., 1964.
2. Velisov E.A. Charles darwin. Evoluutioopin perustajan elämä, työ ja teokset. - M., 1959.
3. Danilova V.S., Kozhevnikov N.N. Luonnontieteen peruskäsitteet. - M .: Aspect Press, 2000 .-- 256 s.
4. Dvorjanski F.A. Darwinismi. - M .: Moskovan valtionyliopisto, 1964 .-- 234 s.
5. Lemeza N.A., Kamlyuk L.V., Lisov ND Käsikirja yliopistoihin tuleville. - M .: Rolf, Iris-press, 1998 .-- 496 s.
6. Mamontov S.G. Biologia: opas yliopiston hakijoille. -M.: valmistua koulusta, 1992 .-- 245 s.
7. Ruzavin G.I. Modernin luonnontieteen käsitteet: Luentokurssi. - M .: Projekti, 2002 .-- 336 s.
8. Sadokhin A.P. Modernin luonnontieteen käsitteet. - M., 2005.
9. Slopov E.F. Modernin luonnontieteen käsitteet. - M .: Vlados, 1999 .-- 232 s.
10. Smygina S.I. Modernin luonnontieteen käsitteet. - Rostov n / a., 1997.




Jotkut hiukkaset siirtyivät vanhemmilta jälkeläisille. Nyt kutsumme näitä hiukkasia geeneiksi. Ajatus korpuskulaarisesta periytymisestä on olennainen ymmärtääksesi, kuinka luonnonvalinta toimii populaatioissa. Evoluutio voidaan nähdä muuttuvana tietyn väestön missä tahansa ominaisuudessa ajan kuluessa. Tietyssä yleisfilosofisessa mielessä tämä on evoluution ydin. ...





Ne pyrittäisiin säilyttämään muuttuneissa olosuhteissa, ja luonnonvalinnan parantamiseen olisi täysi mahdollisuus. 1. LUONNOLLINEN VALINTA ALUEELLISENA EVOLUUTIOTEKIJÄNÄ Kutsuin suotuisten yksilöllisten erojen ja muutosten säilyttämistä ja haitallisten eliminoimista luonnolliseksi valinnaksi tai vahvimman Charles Darwinin kokemukseksi nykyisessä mielessä...
Pienten perinnöllisten muutosten säilyttäminen ja kerääntyminen, joista jokainen on hyödyllinen säilyneelle olennolle. Olosuhteet suotuisat uusien muotojen muodostumiselle luonnonvalinnan kautta. Suuri vaihtelu ja yksilölliset erot ovat tietysti suotuisia olosuhteita. Suuri määrä yksilöitä, mikä lisää mahdollisuuksia kutemaan ...





Siksi niillä on tärkeämpi rooli evoluutiossa. Olennaista on se, että nämä mutaatiot ovat satunnaisia, toisin sanoen niitä ei ole suunnattu. 3. Keskeinen dogmi ja Weismannin periaate hyväksytään. 4. Evoluutio tapahtuu muuttamalla geenien taajuuksia. 5. Nämä muutokset voivat johtua mutaatioista, geenien pääsystä populaatioon ja niiden ulosvirtauksesta siitä, satunnaisesta ajautumisesta ja ...
Perinnöllisyys ja vaihtelevuus ovat organismien ominaisuuksia. Genetiikka tieteenä
Perinnöllisyys- organismien kyky välittää ominaispiirteensä ja kehityspiirteensä jälkeläisilleen.
Vaihtuvuus- erilaisia piirteitä tietyn lajin edustajien keskuudessa sekä jälkeläisten kyky saada eroja vanhempien muodoista.
Genetiikka- tiede perinnöllisyyden ja vaihtelevuuden laeista.
2. Kuvaile tuntemiesi tutkijoiden panosta genetiikan kehitykseen tieteenä ja täytä taulukko.
Genetiikan kehityksen historia
3. Mitä genetiikan menetelmiä tieteenä tunnet?
Genetiikan päämenetelmä on hybridologinen. Tämä on tiettyjen organismien risteyttämistä ja niiden jälkeläisten analysointia. Tätä menetelmää käytti G. Mendel.
Sukututkimus - sukututkimusten tutkimus. Voit määrittää ominaisuuksien periytymismallit.
Twin - identtisten kaksosten vertailu, jonka avulla voit tutkia modifikaatioiden vaihtelua (genotyypin ja ympäristön vaikutuksen määrittämiseksi lapsen kehitykseen).
Sytogeneettinen - tutkimus kromosomijoukon mikroskoopilla - kromosomien lukumäärä, niiden rakenteelliset ominaisuudet. Mahdollistaa kromosomisairauksien tunnistamisen.
4. Mikä on hybridologisen menetelmän ydin ominaisuuksien periytymisen tutkimiseen?
Hybridologinen menetelmä on yksi genetiikan menetelmistä, tapa tutkia organismin perinnöllisiä ominaisuuksia risteyttämällä se sukulaismuodon kanssa ja sitten analysoimalla jälkeläisten ominaisuuksia.
5. Miksi herneitä voidaan pitää menestyneenä geenitutkimuksen kohteena?
Hernelajit eroavat toisistaan pienellä määrällä hyvin erottuvia piirteitä. Herneet on helppo kasvattaa, Tšekissä ne lisääntyvät useita kertoja vuodessa. Lisäksi herneet ovat luonnossa itsepölyttäjiä, mutta kokeessa itsepölytys estyy helposti ja tutkija voi helposti pölyttää kasvin toisen kasvin yhdellä siitepölyllä.
6. Minkä herneiden ominaisuusparien periytymistä G. Mendel tutki?
Mendel käytti 22 puhdasta hernelinjaa. Näiden linjojen kasveilla oli hyvin selvät erot toisistaan: siementen muoto (pyöreä - ryppyinen); siementen väri (keltainen - vihreä); papujen muoto (sileä - ryppyinen); kukkien järjestely varteen (kainaloinen - apikaalinen); kasvin korkeus (normaali - kääpiö).
7. Mitä genetiikassa ymmärretään puhtaana linjana?
Puhdas linja genetiikassa on ryhmä organismeja, joilla on joitakin ominaisuuksia, jotka siirtyvät täysin jälkeläisiin kaikkien yksilöiden geneettisen homogeenisuuden vuoksi.
Perinnön säännöllisyydet. Monohybridiristeys
1. Anna käsitteiden määritelmät.
Alleeliset geenit- geenit, jotka vastaavat yhden piirteen ilmentymisestä.
Homotsygoottinen organismi- organismi, joka sisältää kaksi identtistä alleelista geeniä.
Heterotsygoottinen organismi- organismi, joka sisältää kaksi erilaista alleelista geeniä.
2. Mitä monohybridiristeytys tarkoittaa?
Monohybridiristeytys - risteytys muotoihin, jotka eroavat toisistaan yhdellä vaihtoehtoisten piirteiden parilla.
3. Muotoile tasaisuussääntö ensimmäisen sukupolven hybrideille.
Kun risteytetään kaksi homotsygoottista organismia, jotka eroavat toisistaan yhdellä ominaisuudella, kaikilla ensimmäisen sukupolven hybrideillä on toisen vanhemman piirre ja tämän ominaisuuden sukupolvi on yhtenäinen.
4. Muotoile jakosääntö.
Kun kaksi ensimmäisen sukupolven jälkeläistä (hybridiä) risteytetään toistensa kanssa, toisessa sukupolvessa havaitaan jakautumista ja yksilöitä, joilla on resessiiviset luonteet, ilmaantuu uudelleen; nämä yksilöt muodostavat ¼ osan ensimmäisen sukupolven jälkeläisten kokonaismäärästä.
5. Muotoile sukusolujen puhtauden laki.
Muodostumisen aikana jokainen heistä saa vain yhden kahdesta "perinnöllisyyden elementistä", joka on vastuussa tästä ominaisuudesta.
6. Laadi monohybridiristeyskaavio yleisesti hyväksyttyjen käytäntöjen mukaisesti.

Kuvaa monohybridiristeytymisen sytologisia perusteita käyttämällä tätä esimerkkiä.
P on vanhempien sukupolvi, F1 on jälkeläisten ensimmäinen sukupolvi, F2 on jälkeläisten toinen sukupolvi, A on hallitsevasta piirteestä vastuussa oleva geeni ja on geeni, joka vastaa resessiivisestä ominaisuudesta.
Vanhemman yksilöiden sukusolujen meioosin seurauksena läsnä on yksi geeni, joka on vastuussa tietyn ominaisuuden (A tai a) periytymisestä. Ensimmäisessä sukupolvessa somaattiset solut ovat heterotsygoottisia (Aa), joten puolet ensimmäisen sukupolven sukusoluista sisältää geenin A ja toinen puoli geenin a. Toisen sukupolven sukusolujen satunnaisten yhdistelmien seurauksena syntyy seuraavat yhdistelmät: AA, Aa, aA, aa. Yksilöillä, joilla on kolme ensimmäistä geeniyhdistelmää, on sama fenotyyppi (johtuen hallitsevan geenin läsnäolosta), ja neljännellä erilainen (resessiivinen).
7. Ratkaise geneettinen ongelma monohybridiristeytykselle.
Tavoite 1.
Vesimelonissa hedelmän vihreä väri hallitsee raidallista. Vihreähedelmäisen lajikkeen ja raidallisen lajikkeen risteyttämisestä saatiin ensimmäisen sukupolven hybridejä, joissa oli vihreitä hedelmiä. Hybridit pölytettiin uudelleen ja ne saivat 172 toisen sukupolven hybridiä. 1) Kuinka monta sukusolutyyppiä vihreä hedelmäinen kasvi muodostaa? 2) Kuinka monta F2-kasvia on heterotsygoottisia? 3) Kuinka monta eri genotyyppiä F2:ssa on? 4) Kuinka monta kasvia, joissa on raidallinen hedelmäväri, on F2:ssa? 5) Kuinka monta homotsygoottista kasvia, joissa on vihreitä hedelmiä, on F2:ssa?
Ratkaisu
A - vihreä väri ja - raidallinen väri.
Koska vihreitä ja raidallisia hedelmiä sisältäviä kasveja risteyttäessä saatiin kasveja, joissa oli vihreä hedelmä, voidaan päätellä, että emoyksilöt olivat homotsygoottisia (AA ja aa) (Mendelin ensimmäisen sukupolven hybridien yhtenäisyyssäännön mukaan).
Tehdään risteyskaavio.

Vastaukset:
1,1 tai 2 (heterotsygootin tapauksessa)
2. 86
3. 3
4. 43
5. 43.
Tavoite 2.
Kissojen pitkät hiukset ovat resessiivisiä lyhyille hiuksille. Pitkäkarvainen kissa, risteytetty heterotsygoottisen lyhytkarvaisen kissan kanssa, tuotti 8 pentua. 1) Kuinka monta sukusolutyyppiä kissalla on? 2) Kuinka monta sukusolutyyppiä kissalla on? 3) Kuinka monta fenotyyppisesti erilaista pentua pentueessa on? 4) Kuinka monta genotyypillisesti erilaista pentua pentueessa on? 5) Kuinka monta pitkäkarvaista kissanpentua on?
Ratkaisu
A - lyhyet hiukset, a - pitkät hiukset. Koska kissalla oli pitkä karva, se on homotsygoottinen, hänen genotyyppinsä on aa. Kissalla on genotyyppi Aa (heterotsygoottinen, lyhytkarvainen).
Tehdään risteyskaavio.

Vastaukset:
1. 2
2. 1
3,4 pitkä ja 4 lyhyt
4.4 genotyypin Aa kanssa ja 4 genotyypin aa kanssa
5. 4.
Useita alleeleja. Analysoi ristiä
1. Anna käsitteiden määritelmät.
Fenotyyppi- joukko organismin kaikkia merkkejä ja ominaisuuksia, jotka paljastuvat yksilöllisen kehityksen prosessissa näissä olosuhteissa ja ovat seurausta genotyypin vuorovaikutuksesta sisäisen ja ulkoisen ympäristön tekijöiden kompleksin kanssa.
Genotyyppi- Tämä on joukko organismin kaikkia geenejä, jotka ovat sen perinnöllinen perusta.
2. Miksi hallitsevien ja resessiivisten geenien käsitteet ovat suhteellisia?
Jonkin ominaisuuden geenillä voi olla muita "tiloja", joita ei voida kutsua hallitsevaksi tai resessiiviseksi. Tämä ilmiö voi ilmetä mutaatioiden seurauksena, ja sitä kutsutaan "multiple alleelismiksi".
3. Mitä monialleelismilla tarkoitetaan?
Moninkertainen alleelismi on tietyn geenin useamman kuin kahden alleelin esiintyminen populaatiossa.
4. Täytä taulukko.
Alleelisten geenien vuorovaikutus

5. Mitä on ristin analysointi ja mikä on sen käytännön merkitys?
Analyyttistä risteyttämistä käytetään sellaisten yksilöiden genotyypin määrittämiseen, jotka eivät eroa fenotyypin suhteen. Tässä tapauksessa yksilö, jonka genotyyppi on määritettävä, risteytetään resessiivisen geenin (aa) suhteen homotsygoottisen yksilön kanssa.
6. Ratkaise analysoitavan risteyksen tehtävä.
Tehtävä.
Phlox-terän valkoinen väri hallitsee vaaleanpunaista. Kasvi, jolla on valkoinen teriö, risteytetään kasvin kanssa, jolla on vaaleanpunainen väri. Hybridikasveja saatiin 96, joista 51 valkoista ja 45 vaaleanpunaista. 1) Mitä genotyyppejä emokasveilla on? 2) Kuinka monta sukusolutyyppiä kasvi, jolla on valkoinen teriö, voi muodostaa? 3) Kuinka monta sukusolutyyppiä voi muodostua kasvi, jossa on vaaleanpunainen teriö? 4) Millaista fenotyyppisuhdetta voidaan odottaa F2-sukupolvessa valkokukkaisten F1-hybridikasvien risteyttämisestä?
Ratkaisu.
A on valkoinen väri, a on vaaleanpunainen väri. Yhden kasvin genotyyppi A... on valkoinen, toisen kasvin genotyyppi on vaaleanpunainen.
Koska ensimmäisessä sukupolvessa on jakautuminen 1:1 (51:45), ensimmäisen kasvin genotyyppi on Aa.
Tehdään risteyskaavio.

Vastaukset:
1. Aa ja aa.
2. 2
3. 1
4. 3 valkoisella vispilällä: 1 vaaleanpunaisella vispilällä.
Dihybridiristeys
1. Anna käsitteiden määritelmät.
Dihybridiristeys- yksilöiden risteyttäminen, jotka ottavat erot huomioon kahdella tavalla.
Pennett Lattice Onko englantilaisen geneetikko Reginald Pennettin ehdottama taulukko graafiseksi merkintätyökaluksi vanhempien genotyypeistä peräisin olevien alleelien yhteensopivuuden määrittämiseksi.
2. Mikä on diheterotsygoottien dihybridiristeytyksellä saatujen fenotyyppien suhde? Havainnollista vastausta maalaamalla Punett-ruudukko.
A - Siementen keltainen väri
a - Siementen vihreä väri
B - Tasainen siemenmuoto
c - Siementen ryppyinen muoto.
Sileä keltainen (AABB) × Vihreä ryppyinen (aavv) =
R: AaBb × AaBb (diheterotsygootit)
Sukusolut: AB, Av, AB, AB.
F1 taulukossa:

Vastaus: 9 (keltainen sileä): 3 (vihreä sileä): 3 (keltainen ryppyinen): 1 (vihreä ryppyinen).
3. Muotoile ominaisuuksien itsenäisen periytymisen laki.
Dihybridiristeytyksissä geenit ja ominaisuudet, joista nämä geenit ovat vastuussa, periytyvät toisistaan riippumatta.
4. Ratkaise geneettiset ongelmat dihybridiristeyttämiseen.
Tavoite 1.
Kissojen musta väri hallitsee kellanruskeaa ja lyhyt karva pitkiä. Puhdasrotuiset persialaiset kissat (musta pitkäkarvainen) risteytettiin siamilaisten kanssa (fawn lyhytkarvainen). Tuloksena saadut hybridit risteytyivät keskenään. Mikä on todennäköisyys saada puhdasrotuinen siamilainen kissanpentu F2:ssa? kissanpentu, joka on fenotyyppisesti samanlainen kuin persialainen; pitkäkarvainen fawn kissanpentu (ilmaista osissa)?
Ratkaisu:
A - väri musta ja - kellanruskea.
B - lyhyet hiukset, c - pitkät.

Tehdään Punnet-hila.

Vastaus:
1) 1/16
2) 3/16
3) 1/16.
Tavoite 2.
Tomaatissa hedelmän pyöreä muoto hallitsee päärynän muotoista ja hedelmän punainen väri hallitsee keltaista. Heterotsygoottisen, punaisen väriltään ja päärynän muotoisen hedelmäisen kasvin ja pyöreähedelmäisen keltahedelmäisen kasvin risteyttämisestä saatiin 120 kasvia. 1) Kuinka monta sukusolutyyppiä heterotsygoottinen kasvi, jolla on punainen hedelmäväri ja päärynän muotoinen muoto, muodostuu? 2) Kuinka monta eri fenotyyppiä saat tästä risteyksestä? 3) Kuinka monta eri genotyyppiä saat tästä risteyksestä? 4) Kuinka monta kasvia, joilla oli punainen väri ja pyöreä hedelmämuoto, tuli? 5) Kuinka monella kasvilla oli keltainen väri ja pyöreä hedelmämuoto?
Ratkaisu
A - pyöristetty muoto ja - päärynän muotoinen.
B - punainen väri, in - keltainen väri.
Selvitämme vanhempien genotyypit, sukusolujen tyypit ja kirjoitamme risteytyskaavion.

Tehdään Punnet-hila.

Vastaus:
1. 2
2. 4
3. 4
4. 30
5. 30.
Perinnöllisyyden kromosomiteoria. Nykyaikaiset käsitykset geenistä ja genomista
1. Anna käsitteiden määritelmät.
Crossover- homologisten kromosomien alueiden vaihtoprosessi konjugaation aikana meioosin I-vaiheessa.
Kromosomi kartta- Tämä on kaavio tiettyjen samassa kytkentäryhmässä olevien kromosomien geenien keskinäisestä järjestelystä ja suhteellisista etäisyyksistä.
2. Milloin piirteiden itsenäisen periytymisen lakia rikotaan?
Ylityksessä Morganin lakia rikotaan, eivätkä yhden kromosomin geenit periydy, koska osa niistä korvataan homologisen kromosomin alleelisilla geeneillä.
3. Kirjoita T. Morganin perinnöllisyyden kromosomiteorian päämääräykset.
Geeni on osa kromosomista.
Alleeliset geenit (yhdestä ominaisuudesta vastaavat geenit) sijaitsevat homologisten kromosomien tiukasti määritellyissä paikoissa (lokuksissa).
Geenit sijaitsevat kromosomeissa lineaarisesti eli peräkkäin.
Sukusolujen muodostumisprosessissa homologisten kromosomien välillä tapahtuu konjugaatiota, jonka seurauksena ne voivat vaihtaa alleelisia geenejä, eli risteytys voi tapahtua.
4. Muotoile Morganin laki.
Geenit, jotka sijaitsevat samassa kromosomissa, putoavat meioosin aikana yhdeksi sukusoluksi, eli ovat periytyviä.
5. Mikä määrittää kahden ei-alleelisen geenin eron todennäköisyyden risteytyksissä?
Kahden ei-alleelisen geenin eron todennäköisyys risteytyksen aikana riippuu niiden välisestä etäisyydestä kromosomissa.
6. Mihin organismien geneettinen kartoitus perustuu?
Laskemalla risteytymistiheyden minkä tahansa kahden saman kromosomin geenin välillä, jotka ovat vastuussa erilaisista ominaisuuksista, voidaan määrittää tarkasti näiden geenien välinen etäisyys ja siksi alkaa rakentaa geneettistä karttaa, joka on kaavio suhteellisesta sijainnista. geeneistä, jotka muodostavat yhden kromosomin.
7. Mitä varten kromosomikartat ovat?
Geenikarttojen avulla saat selville eläinten ja kasvien geenien sijainnin ja tietoa niistä. Tämä auttaa taistelussa erilaisia parantumattomia sairauksia vastaan.
Perinnöllinen ja ei-perinnöllinen vaihtelu
1. Anna käsitteiden määritelmät.
Reaktionopeus- genotyypin kyky muodostua ontogeneesissä, riippuen ympäristöolosuhteista, erilaisista fenotyypeistä. Se luonnehtii ympäristön osuutta ominaisuuden toteutumisessa ja määrittelee lajin modifikaatiovaihteluita.
Mutaatio- pysyvä (eli sellainen, jonka tietyn solun tai organismin jälkeläiset voivat periä) genotyypin transformaatio, joka tapahtuu ulkoisen tai sisäisen ympäristön vaikutuksesta.
2. Täytä taulukko.

3. Mikä määrittää modifikaatiovaihteluiden rajat?
Modifikaatiovaihtelurajat riippuvat reaktionopeudesta, joka määräytyy geneettisesti ja periytyy.
4. Mitä yhteistä on kombinatiivisella ja mutaatiovaihtelulla ja miten ne eroavat toisistaan?
Yleistä: Molemmat vaihtelutyypit johtuvat geneettisen materiaalin muutoksista.
Erot: kombinatiivinen vaihtelevuus johtuu geenien rekombinaatiosta sukusolujen fuusion aikana, ja mutaatioiden vaihtelu johtuu mutageenien vaikutuksesta kehoon.
5. Täytä taulukko.
Mutaatioiden tyypit

6. Mitä mutageeniset tekijät tarkoittavat? Anna asiaankuuluvia esimerkkejä.
Mutageeniset tekijät ovat vaikuttajia, jotka johtavat mutaatioiden esiintymiseen.
Nämä voivat olla fysikaalisia vaikutuksia: ionisoiva säteily ja ultraviolettisäteily vahingoittavat DNA-molekyylejä; kemikaalit, jotka häiritsevät DNA:n rakenteita ja replikaatioprosesseja; virukset, jotka lisäävät geeninsä isäntäsolun DNA:han.
Ominaisuuksien periytyminen ihmisiin. Perinnölliset sairaudet ihmisillä
1. Anna käsitteiden määritelmät.
Geenisairaudet- geeni- tai kromosomimutaatioiden aiheuttamat sairaudet.
Kromosomitaudit- kromosomien lukumäärän tai niiden rakenteen muutoksesta johtuvat sairaudet.
2. Täytä taulukko.
Ominaisuuksien periytyminen ihmisiin


3. Mitä sukupuoleen liittyvällä perinnöllä tarkoitetaan?
Sukupuolisidonnainen perinnöllisyys on sellaisten ominaisuuksien periytymistä, joiden geenit sijaitsevat sukupuolikromosomeissa.
4. Mitkä ominaisuudet ihmisissä ovat sukupuolisidonnaisia?
Hemofilia ja värisokeus ovat periytyviä ihmisillä.
5. Ratkaise geneettisiä ongelmia ominaisuuksien periytymiseksi ihmisillä, mukaan lukien sukupuoleen liittyvä perinnöllisyys.
Tavoite 1.
Ihmisillä pitkien ripsien geeni hallitsee lyhyiden ripsien geeniä. Nainen, jolla oli pitkät silmäripset, jonka isällä oli lyhyet silmäripset, naimisissa miehiä, joilla oli lyhyet silmäripset. 1) Kuinka monta sukusolutyyppiä naisella on? 2) Kuinka monen tyyppisiä sukusoluja muodostuu miehillä? 3) Millä todennäköisyydellä tässä perheessä syntyy pitkäsilmäinen lapsi (%)? 4) Kuinka monta eri genotyyppiä ja kuinka monta fenotyyppiä voi olla tietyn avioparin lapsilla?
Ratkaisu
A - pitkät ripset
a - lyhyet ripset.
Naiset ovat heterotsygoottisia (Aa), koska isällä oli lyhyet silmäripset.
Mies on homotsygoottinen (aa).

Vastaus:
1. 2
2. 1
3. 50
4. 2 genotyyppiä (Aa) ja 2 fenotyyppiä (pitkät ja lyhyet ripset).
Tavoite 2.
Ihmisillä vapaa korvalehti hallitsee ei-vapaata, ja sileä leuka on resessiivinen suhteessa leukaan, jossa on kolmion muotoinen kuoppa. Nämä ominaisuudet periytyvät itsenäisesti. Miehen, jolla oli ei-vapaa korvalehti ja kolmion muotoinen kuoppa leuassa, ja naisen, jolla oli vapaa korvalehti ja sileä leuka, avioliitosta syntyi poika, jolla oli sileä leuka ja ei-vapaa korvalehti. Mikä on todennäköisyys, että tässä perheessä syntyy lapsi, jolla on sileä leuka ja vapaa korvalehti? jossa on kolmion muotoinen kuoppa leuassa (%)?
Ratkaisu
A - vapaa korvalehti
a - ei-vapaa korvalehti
B - kolmion muotoinen kuoppa
c - sileä leuka.
Koska parilla oli lapsi, jolla on homotsygoottisia piirteitä (aavv), äidin genotyyppi on Aavb ja isän aaBb.
Kirjataan ylös vanhempien genotyypit, sukusolujen tyypit ja risteytyskaavio.

Tehdään Punnet-hila.

Vastaus:
1. 25
2. 50.
Tavoite 3.
Ihmisillä hemofiliaa aiheuttava geeni on resessiivinen ja sijaitsee X-kromosomissa, ja albinismi johtuu autosomaalisesta resessiivisestä geenistä. Vanhemmilla, jotka olivat normaaleja näiltä ominaisuuksiltaan, oli albiino- ja hemofiilipoika. 1) Mikä on todennäköisyys, että heidän seuraava poikansa osoittaa nämä kaksi epänormaalia merkkiä? 2) Mikä on todennäköisyys saada terveitä tyttäriä?
Ratkaisu:
X ° - hemofilian esiintyminen (resessiivinen), X - hemofilian puuttuminen.
A - normaali ihonväri
a - albiino.
Vanhempien genotyypit:
Äiti - X ° HAa
Isä - HUAa.
Tehdään Punnet-hila.

Vastaus: albinismin ja hemofilian (genotyyppi X ° Waa) merkkien ilmenemisen todennäköisyys - seuraavassa pojassa - 6,25%. Todennäköisyys saada terveitä tyttäriä - (genotyyppi XXAA) - 6,25%.
Tehtävä 4.
Ihmisten verenpaineen määrää hallitseva autosomaalinen geeni, ja optisen atrofian aiheuttaa resessiivinen sukupuoleen liittyvä geeni. Näköatrofiasta kärsinyt nainen meni naimisiin verenpainetautia sairastavan miehen kanssa, jonka isä oli myös verenpainetauti ja äiti terve. 1) Millä todennäköisyydellä tämän perheen lapsi kärsii molemmista poikkeavuuksista (%)? 2) Mikä on todennäköisyys saada terve vauva (%)?
Ratkaisu.
X ° - atrofian esiintyminen (resessiivinen), X - atrofian puuttuminen.
A - verenpainetauti
a - ei hypertensiota.
Vanhempien genotyypit:
Äiti - X ° X ° aa (koska hän on sairas atrofiasta ja ilman verenpainetautia)
Isä - HUAa (koska hän ei ole sairas atrofiasta ja hänen isänsä sairastui verenpaineeseen ja hänen äitinsä on terve).
Tehdään Punnet-hila.

Vastaus:
1. 25
2. 0 (vain 25 %:lla tyttäristä ei ole näitä puutteita, mutta he ovat atrofian kantajia ja ilman verenpainetautia).
Perinnöllisyys - Tämä on elävien organismien ominaisuus säilyttää ja siirtää ominaisuuksia useiden sukupolvien ajan. Perinnöllisyydestä johtuen lajin, rodun ominaisuudet säilyvät sukupolvelta toiselle.
Perinnöllinen variaatio (mutaatio tai genotyyppi) liittyy yksilön genotyypin muutokseen, joten tuloksena olevat muutokset periytyvät. Hän on luonnonvalinnan materiaali. Darwin kutsui tätä perintöä määrittelemättömäksi. Perinnöllisen vaihtelun perustana ovat mutaatiot - äkilliset äkilliset ja suuntaamattomat muutokset alkuperäisessä muodossa. Ne johtavat eläviin organismeihin laadullisesti uusien perinnöllisten ominaisuuksien ja ominaisuuksien ilmaantumiseen, joita ei aiemmin ollut luonnossa. Perinnöllisen vaihtelun lähde on mutaatioprosessi. Mutaatioita on useita: genominen, kromosomaalinen ja geenimutaatio.
Genomimutaatiot (polyploidia ja aneuploidia) - Nämä ovat muutoksia kromosomien lukumäärässä. Polyploidia on haploidisten kromosomien (Zn, 4n jne.) moninkertainen lisääntyminen. Useimmiten polyploidia muodostuu, kun kromosomien eroaminen solun napoihin rikkoutuu meioosissa tai mitoosissa mutageenisten tekijöiden vaikutuksesta. Se on laajalle levinnyt kasveissa ja erittäin harvinainen eläimissä.
Aneuploidia - kromosomien lukumäärän lisääntyminen tai väheneminen yksittäisissä pareissa. Se tapahtuu, kun kromosomit eivät eroa meioosissa tai kromatidit mitoosissa. Aneuploideja löytyy kasveista ja eläimistä, ja niille on ominaista alhainen elinkelpoisuus.
Kromosomimutaatiot - Nämä ovat muutoksia kromosomien rakenteessa. On olemassa seuraavan tyyppisiä kromosomimutaatioita:
Defiration - kromosomien pääteosien menetys.
Poistot - kromosomien olkapään osan menetys.
Monistaminen - geenisarjan toistuminen tietyssä kromosomin osassa.
Käännös - kromosomiosan kierto 180°.
Translokaatio - kohdan siirtyminen saman kromosomin toiseen päähän tai toiseen, ei-homologiseen kromosomiin.
Geenimutaatiot - muutokset DNA-molekyylin (geenin) nukleotidisekvenssissä. Niiden seurauksena polypeptidiketjun aminohappojen sekvenssi muuttuu ja proteiinin ilmaantuminen uusilla ominaisuuksilla. Useimmat geenimutaatiot eivät ilmene fenotyyppisesti, koska ne ovat resessiivisiä.
Sytoplasmiset mutaatiot - liittyy muutoksiin DNA:ta sisältävissä sytoplasmisissa organelleissa (mitokondriot ja plastidit). Nämä mutaatiot ovat äidin kautta periytyviä, koska tsygootti vastaanottaa opsn-additiossa koko sytoplasman munasolusta. Esimerkki: Kasvien kirjavaisuus liittyy kloorilastien mutaatioihin.
Merkitys evoluutiossa ja ontogeneesissä Sukusoluihin vaikuttavat mutaatiot (generatiiviset mutaatiot) ilmestyvät seuraavassa sukupolvessa. Somaattisten solujen mutaatioita esiintyy niissä elimissä, jotka sisältävät muuttuneet solut. Eläimillä somaattiset mutaatiot eivät periydy, koska uusi organismi ei synny somaattisista soluista. Kasveissa, jotka lisääntyvät vegetatiivisesti, somaattiset mutaatiot voivat jatkua. Mutaatiovariaatiolla on evoluution perinnöllisten muutosten päätoimittaja. Hän on kaikkien evolutionaaristen muutosten ensisijainen materiaali.
Genotyyppinen vaihtelu ja sen tyypit. Merkitys ontogeniassa ja evoluutiossa.
Genotyyppinen tai perinnöllinen vaihtelu, on fenotyypin muutos, joka johtuu genotyypin muutoksista.
Se johtuu mutaatioista ja niiden yhdistelmistä seksuaalisen lisääntymisen aikana (esimerkiksi karjan perinnöllinen sarvettomuus).
Perinnöllinen ja kombinatiivinen vaihtelu erotetaan geneettisen materiaalin vaihtelun luonteesta riippuen. Kombinatiivinen vaihtelevuus johtuu uusien genotyyppien geeniyhdistelmien muodostumisesta jälkeläisissä, jotka muodostuvat geenien ja kromosomien rekombinaatioiden seurauksena seksuaalisen lisääntymisen aikana. Elävien organismien genotyyppien ääretön valikoima, kunkin genotyypin ainutlaatuisuus johtuu kombinatiivisesta vaihtelevuudesta. Tämän tyyppisellä vaihtelulla geenien yhdistelmät ja niiden vuorovaikutuksen luonne genotyypissä muuttuvat, kun taas geenit itse pysyvät ennallaan.
Kombinatiivinen vaihtelu , ovat seurausta vanhempien geenien rekombinaatiosta jälkeläisten genotyypeissä, perustuu kolmeen päämekanismiin.
1. Itsenäinen hajoaminen tytärsoluihin (spermatosyytit II, munasolu II ja ensimmäinen pelkistyskappale) homologiset kromosomit kustakin parista (tapahtuu meioosin I jakautumisen aikana gametogeneesin aikana). Esimerkiksi jopa 2 kromosomiparille on mahdollista 2 muunnelmaa kromosomien hajoamisesta tytärsoluiksi ja 4 siittiötyyppiä (kuva 76).
2. Satunnainen sukusolujen yhdistelmä, ja siksi homologiset (isän ja äidin) kromosomit hedelmöityksen aikana. Edellä mainituissa neljässä siittiötyypissä yhden niistä osallistuminen munasolun hedelmöitykseen on täysin satunnaista, ja tulokset yhden miehen kromosomien muunnelman spesifisestä yhdistelmästä yhden kanssa (myös neljästä mahdollisesta, koska pelkistyskappaleet kuljettavat pois kolme varianttia ja lakkasivat olemasta ) niille homologisista naiskromosomien muunnelmista.
3. Yksittäisten alleelien vaihto homologisten kromosomien välillä meioosin ylityksen aikana. Sen jälkeen siittiöiden kromosomien alleeliyhdistelmille on tunnusomaista uudet variantit, jotka poikkeavat kehon somaattisten solujen vastaavista (kuva 77).
Crossover tapahtuu meioosin alussa, kun homologiset kromosomit asettuvat toisiaan vastaan. Tässä tapauksessa homologisten kromosomien osat leikkaavat, katkeavat ja kiinnittyvät sitten uudelleen, mutta tällä kertaa toiseen kromosomiin. Lopulta muodostuu neljä kromosomia erilaisilla geeniyhdistelmillä. Kromosomit, joita kutsutaan "rekombinanttikromosomeiksi", sisältävät uusia geeniyhdistelmiä (Ab ja ab), jotka puuttuivat alkuperäisistä kromosomeista (AB ja ab).
Kombinatiivinen vaihtelevuus selittää, miksi lapsista löytyy uusia äidin ja isän sukulaisten piirteiden yhdistelmiä ja sellaisia erityisiä muunnelmia, jotka eivät olleet ominaisia isälle, äidille, isoisälle tai isoäidille jne.
Kombinatiivisen vaihtelevuuden ansiosta jälkeläisissä syntyy erilaisia genotyyppejä, millä on suuri merkitys evoluutioprosessille johtuen siitä, että: 1) evoluutioprosessin materiaalin monimuotoisuus lisääntyy heikentämättä yksilöiden elinkykyä; 2) organismien mahdollisuudet sopeutua muuttuviin ympäristöolosuhteisiin laajenevat ja siten varmistetaan organismiryhmän (populaation, lajin) selviytyminen kokonaisuutena.
Jalostuksessa käytetään kombinatiivista vaihtelua taloudellisesti arvokkaamman perinnöllisten ominaisuuksien yhdistelmän saamiseksi. Erityisesti käytetään heteroosilmiötä, elinkelpoisuuden lisääntymistä, kasvun intensiteettiä ja muita indikaattoreita hybridisaation aikana eri alalajien tai lajikkeiden edustajien välillä. Ilmiö antaa päinvastaisen vaikutuksensukusiitos tai siihen läheisesti liittyvä risteytys - organismien risteyttäminen yhteisten esi-isiensä kanssa. Ristikkäisten organismien yhteinen alkuperä lisää niiden todennäköisyyttä, että niillä on samat alleelit minkä tahansa geenin kanssa, ja siten homotsygoottisten organismien ilmaantumisen todennäköisyyttä. Suurin sisäsiitosaste saavutetaan kasveilla itsepölytyksissä ja eläimillä itsehedelmöityksessä. Homotsygoottisuus lisää resessiivisten alleelisten geenien ilmenemismahdollisuutta, joiden mutageeniset muutokset johtavat perinnöllisistä poikkeavuuksista kärsivien organismien ilmaantumiseen.
Kombinatiivisen vaihtelevuuden ilmiön tutkimuksen tuloksia hyödynnetään lääketieteellisessä ja geneettisessä neuvonnassa, erityisesti sen toisessa ja kolmannessa vaiheessa: jälkeläisten ennustamisessa, päätelmän muodostamisessa ja geneettisen riskin merkityksen selityksessä.
Avioliittojärjestelmien ohella erotetaan kaksi avioparien muodostumistyyppiä:
1) avioparien positiivinen assortatiivinen (valikoiva) muodostuminen tai tietyiltä fenotyyppisiltä ominaisuuksiltaan samankaltaisten yksilöiden tiheämpi avioliitto (avioliitot kuurojen ja tyhmien tai samanpituisten, henkisesti kehittyneiden välillä jne.);
2) avioparien negatiivinen assortatiivinen muodostuminen tai harvinaisempi avioliitto henkilöiden, joilla on samanlaiset tietyt ominaisuudet (esim. punatukkaiset välttelevät avioliittoa).
Sekä sisäsiitos että paritteluparien positiivinen assortatiivinen muodostuminen lisäävät (jälkimmäinen, joskin vähäisemmässä määrin) jälkeläisten homotsygoottisuuden tasoa, myös haitallisten resessiivisten alleelien paikoissa. Sitä vastoin ulkosiitos lisää heterotsygoottisuuden astetta ja monissa tapauksissa lisää elinkelpoisuuden tasoa. Sukusiitoksen ja avioparien positiivisen assortatiivisen muodostumisen mahdollisia seurauksia hyödynnetään mahdollisten aviopuolisoiden lääketieteellisessä geneettisessä neuvonnassa.
Mutaatiot - Nämä ovat perinnöllisiä muutoksia geneettisessä materiaalissa, jotka johtavat muutokseen organismin ominaisuuksissa. Mutaatioteorian perustan loi G. de Vries jo vuonna 1901, joka kuvasi mutaatioita eloterassa, mutta niiden molekyylimekanismeja tutkittiin paljon myöhemmin. G. de Vriesin mukaan mutaatio on äkillinen, ajoittainen muutos perinnöllisissä piirteissä.
H. de Vriesin mutaatioteorian ydin rajoittuu seuraaviin säännöksiin:
1) mutaatio tapahtuu diskreetti, ilman siirtymiä;
2) uudet muodot ovat vakioita;
3) mutaatiot ovat monisuuntaisia (hyödyllisiä ja haitallisia);
4) mutaatioiden havaitsemisnopeus riippuu tutkittavien organismien näytteen koosta;
5) samat mutaatiot voivat esiintyä toistuvasti.
Mutaatiomuutokset ovat erittäin vaihtelevia. Ne voivat vaikuttaa lähes kaikkiin organismin morfologisiin, fysiologisiin ja biokemiallisiin ominaisuuksiin, voivat aiheuttaa teräviä tai päinvastoin tuskin havaittavia fenotyyppisiä poikkeamia normista.
Mutaatiovaihtelu perustuu geenien ja kromosomien rakenteellisiin muutoksiin. Geneettisen materiaalin muutosten luonteesta riippuen on olemassa:
1) geeni(piste)mutaatiot, jotka ovat nukleotidiparin lisäystä, katoamista, korvaamista tai muutosta;
2) insertio - DNA-molekyylien tai niiden fragmenttien liittäminen ("leikkaus") geeniin, mikä useimmiten johtaa sen inaktivoitumiseen tai voimakkaaseen polaariseen vaikutukseen operoneissa;
3) kromosomien uudelleenjärjestelyt tai poikkeamat - kromosomien rakenteen muutokset niiden repeämisen perusteella;
4) genomiset (genotyyppiset) mutaatiot, jotka muodostuvat solun kromosomien lukumäärän muutoksesta.
Fenotyyppinen vaihtelu ja sen tyypit. Muutosten mukautuva luonne. Ominaisuuden reaktionopeus. Ominaisuuden ilmaisukyky ja tunkeutuminen.
Modifikaatio (fenotyyppinen) vaihtelu johtuu vain ulkoisten olosuhteiden vaikutuksesta, eikä se liity genotyypin muutokseen. Fenotyypin tilan tiettyjä muunnelmia, joissa on modifikaatiovaihtelua, kutsutaan modifikaatioiksi. Suurin kiinnostus ovatmukautuvat muutokset - ei-perinnölliset muutokset, jotka ovat hyödyllisiä keholle ja edistävät sen selviytymistä muuttuneissa olosuhteissa. Toisin kuin mutaatiot (harvinaiset, yksittäiset ja satunnaiset tapahtumat), mukautuvat modifikaatiot ovat suunnattuja ja samalla usein palautuvia, ennustettavia ja usein ominaisia suurille organismiryhmille. Modifikaatioiden olemassaolo perustuu siihen, että fenotyyppi on genotyypin ja ulkoisten olosuhteiden vuorovaikutuksen tulos. Siksi muutos ulkoisissa olosuhteissa voi aiheuttaa muutoksia fenotyypissä, joihin ei liity muutoksia genotyypissä. Muutosten ilmaantumisen mekanismi on siinä, että ympäristöolosuhteet vaikuttavat kehittyvässä organismissa tapahtuviin entsymaattisiin reaktioihin (aineenvaihduntaprosesseihin) ja muuttavat jossain määrin niiden kulkua ja siten tulosta - muodostuneen ominaisuuden tilaa. niiden perusteella.
Muutoksilla on seuraavat ominaisuudet:
1) modifikaation vakavuus on verrannollinen modifikaation aiheuttavan tekijän elimistöön kohdistuvan vaikutuksen voimakkuuteen ja kestoon (tämä malli erottaa pohjimmiltaan modifikaatiot mutaatioista, erityisesti geenimutaatioista);
2) suurimmassa osassa tapauksista modifikaatio on kehon hyödyllinen adaptiivinen reaktio vastauksena jonkin ulkoisen tekijän vaikutukseen
3) vain ne muutokset ovat mukautuvia , jotka johtuvat tavanomaisista luonnonolosuhteiden muutoksista, joihin tämän lajin yksilöiden esi-isät ovat toistuvasti "kohdatneet" sen menneen evoluutiohistorian aikana;
4) kokeellisten vaikutusten, erityisesti kemiallisten ja fysikaalisten tekijöiden, aiheuttamat muutokset, joita organismi ei luonnossa kohtaa, ei pääsääntöisesti ole mukautuvaa arvoa ja edustaa usein epämuodostumia ja epämuodostumia. Tällä tavalla indusoituja modifikaatioita kutsutaan usein morfooseiksi.
5), toisin kuin mutaatiot, joille on ominaista korkea pysyvyys, modifikaatioilla on vaihteleva vastustuskyky. Monet niistä ovat palautuvia, ts. syntyneet muutokset häviävät vähitellen, jos ne aiheuttaneen tekijän toiminta lakkaa. Joten ihmisen auringonpolttama katoaa, kun iho lakkaa olemasta alttiina auringolle, lihasten tilavuus vähenee harjoituksen lopettamisen jälkeen jne.
6) modifikaatiot, toisin kuin mutaatiot, eivät ole periytyviä, ts. ovat ei-perinnöllisiä. Tämä on yhdenmukainen "molekyylibiologian keskeisen dogman" F. Crickin kanssa, jonka mukaan tiedon siirto on mahdollista vain geneettisestä materiaalista geenituotteisiin-proteiineihin, mutta ei päinvastaiseen suuntaan.
Ulkoisilla olosuhteilla on valtava vaikutus kehittyvän organismin kaikkiin merkkeihin ja ominaisuuksiin.
Reaktionopeus. Modifikaatiovaihteluilla ominaisuus voi muuttua tietyissä genotyypin kullekin tilassa ominaisissa rajoissa (alueet). Aluetta, jolla sama genotyyppi pystyy määrittämään eri fenotyyppien kehittymisen, kutsutaan reaktionopeudeksi. Toisin sanoen normireaktiot ovat organismin, jolla on spesifinen muuttumaton genotyyppi, ontogeneesin mahdollisen vaihtelun amplitudi. Parhaiten vastenopeus havaitaan organismeissa, joilla on samat genotyypit, esimerkiksi vegetatiivisesti lisääntyvillä kasveilla ja identtisillä kaksosilla. Tässä tapauksessa on mahdollista tunnistaa genotyypin reaktionopeus "puhtaimmassa" muodossa. Genotyypin ohjaama vastenopeus on evoluutioprosessin tulos.
Tärkeimmät tekijät, jotka voivat aikaansaada merkkien vaihtelun normaalin reaktioalueen sisällä, ovat:
1) ominaisuuden polygeeninen määritys ja organismin reaktio;
2) geenin pleiotrooppinen vaikutus;
3) mutaation ilmenemisen riippuvuus ympäristöolosuhteista;
4) organismin heterotsygoottisuus;
5) geenien vuorovaikutus geenituotteiden (proteiinimolekyylien alayksiköiden) tasolla;
6) vaihtoehtoiset kehitystavat kehon järjestelmässä ja biosynteesin toteuttaminen solussa (yhden polun estämistä kompensoi toinen).
Läpäisevyys jolle on tunnusomaista tietyn geenin alleelin ilmentymistiheys tai todennäköisyys, ja se määräytyy populaation yksilöiden prosenttiosuuden perusteella, jossa se ilmeni fenotyyppisesti. Erota täydellinen (piirteen ilmeneminen kaikissa yksilöissä) ja epätäydellinen (osittain) penetranssi. Kvantitatiivisesti penetranssi ilmaistaan yksilöiden prosenttiosuutena prosentteina, joissa tämä alleeli ilmenee. Esimerkiksi synnynnäisen lonkkanivelen tunkeutumisaste ihmisillä on 25 %, mikä osoittaa, että vain 1/4 genotyypeistä, jotka kantavat tiettyä geeniä, osoittavat sen fenotyyppistä vaikutusta.
Taustalla epätäydellinen penetranssi on geneettisten ja ympäristöllisten syiden vuorovaikutus. Tiettyjen alleelien tunkeutumisen tuntemus on välttämätöntä lääketieteellisessä ja geneettisessä neuvonnassa, jotta voidaan määrittää "terveiden" ihmisten mahdollinen genotyyppi, joiden perheessä on esiintynyt perinnöllisiä sairauksia. Epätäydellisen tunkeutumisen tapaukset sisältävät geenien ilmentymisen, jotka säätelevät sukupuolirajoitettuja ja sukupuolesta riippuvia piirteitä.
Ilmaisukyky - geenin fenotyyppisen ilmentymisen aste sen toiminnan voimakkuuden mittana, jonka määrittää piirteen kehitysaste. Ilmeisyys voi molemmilla sukupuolilla olla sama tai erilainen, vakio tai vaihteleva, jos saman genotyypin piirteen vakavuus vaihtelee yksilöittäin. Tämän alleelin hallitseman ominaisuuden vaihtelevuuden puuttuessa puhutaan jatkuvasta ekspressiivisuudesta (yksiselitteisestä reaktionopeudesta). Esimerkiksi ihmisen ABO-veriryhmien alleeleilla on käytännössä jatkuva ekspressio. Toinen ilmeisyyden tyyppi on epävakaa tai vaihteleva. Se perustuu useisiin syihin: ympäristön ulkopuolisten olosuhteiden vaikutus (muunnos), genotyyppinen ympäristö (geenien vuorovaikutuksessa).
Ilmaisukykyä arvioidaan kvantitatiivisesti tilastollisten indikaattoreiden avulla. Ekspressiivyyden muutosten äärimmäisissä varianteissa (piirteen täydellinen puuttuminen) käytetään lisäominaisuutta - penetranssia. Huntingtonin korea voi toimia esimerkkinä hallitsevan geenin epätäydellisestä penetraatiosta ja vaihtelevasta ekspressiivisuudesta. Huntingtonin korean ensimmäisen ilmestymisen ikä vaihtelee. Tiedetään, että joissakin kantajissa se ei koskaan ilmene (epätäydellinen penetranssi); lisäksi tällä geenillä on vaihteleva ekspressio, koska kantajat sairastuvat eri ikäisinä.
Modifikaatiovaihtelu tarjoaa suhteellisen nopean muodostumisen organismin sopeutumisesta muuttuviin ympäristöolosuhteisiin ontogeneaation aikana, mikä myötävaikuttaa organismin selviytymiseen. Näin ollen modifikaatiot ovat tärkein tekijä elävän organismin normaalissa kehityksessä ja ontogeneesin loppuunsaattamisessa.
Huolimatta siitä, että jälkeläiset eivät peri modifikaatioita, modifikaatioiden vaihtelevuus yleensä on tärkeää orgaanisen maailman evoluution kannalta. Luonnollisen valinnan aikana modifikaatiot voivat toimia "peitteenä" mutaatioille, joiden fenotyyppinen ilmentymä monistaa ei-perinnöllisiä muutoksia. Suosittelemalla organismien selviytymistä modifikaatiovaihtelu edistää tiettyjen eri genotyyppien omaavien yksilöiden säilymistä ja lisääntymiseen osallistumista. Tämän lisäksi muutokset edistävät lajien (populaation) uusien elinympäristöjen kehittymistä, mikä johtaa tämän organismiryhmän levinneisyyteen. Kaikki nämä muutosvaikutukset edistävät lajin tai populaation evoluution menestystä.
Ihminen geenitutkimuksen erityisobjektina. Ihmisgenetiikan tutkimusmenetelmät. Avioliiton lääketieteellinen ja geneettinen puoli. Lääketieteellinen geneettinen neuvonta. Genetiikan merkitys lääketieteessä.
Ihminen geenitutkimuksen erityisobjektina. Ihmisgenetiikan tutkimukseen liittyy suuria vaikeuksia: monimutkainen karyotyyppi - monia kromosomeja ja kytkentäryhmiä, myöhäinen murrosikä ja harvinainen sukupolvien vaihto, pieni määrä jälkeläisiä, mahdottomuus kokeilla, mahdottomuus luoda samoja elinoloja. Kaikesta tästä huolimatta ihmisen genetiikka on nykyään paremmin tutkittu kuin monien muiden organismien (esimerkiksi nisäkkäiden) genetiikka lääketieteen tarpeiden ja monien nykyaikaisten tutkimusmenetelmien vuoksi.
Opiskelumenetelmät :
Sukututkimusmenetelmä Se koostuu sukuluetteloiden tutkimisesta Mendelin perintölakien perusteella ja auttaa määrittämään piirteen periytymisen luonteen (dominoiva tai resessiivinen). Näin määritetään henkilön yksilöllisten ominaisuuksien periytyminen: kasvojen piirteet, pituus, veriryhmä, henkinen ja henkinen meikki sekä jotkut sairaudet. Tämä menetelmä paljasti läheisten avioliittojen haitalliset seuraukset, jotka ilmenevät erityisesti homotsygoottisuudessa saman epäsuotuisan resessiivisen alleelin suhteen. Perinnöllisissä avioliitoissa todennäköisyys saada lapsia, joilla on perinnöllisiä sairauksia ja varhainen lapsikuolleisuus, on kymmeniä ja jopa satoja kertoja keskimääräistä suurempi.
Kaksoismenetelmä on tutkia identtisten kaksosten välisiä eroja. Tämä menetelmä on luonnon itsensä tarjoama. Se auttaa tunnistamaan ympäristöolosuhteiden vaikutuksen fenotyyppiin, jolla on samat genotyypit. Samoissa olosuhteissa kasvaneet identtiset kaksoset ovat hämmästyttävän samankaltaisia paitsi morfologisissa ominaisuuksissa, myös henkisissä ja älyllisissä ominaisuuksissa. Kaksoismenetelmä paljasti perinnöllisyyden roolin useissa sairauksissa.
Väestötilastollinen menetelmä. Populaatiogenetiikka tutkii yksittäisten ihmisryhmien (populaatioiden) välisiä geneettisiä eroja, tutkii geenien maantieteellisen jakautumisen malleja.
Sytogeneettinen menetelmä . perustuu vaihtelevuuden ja perinnöllisyyden tutkimukseen solu- ja solurakenteiden tasolla. Useiden vakavien sairauksien ja kromosomien poikkeavuuksien välillä on havaittu yhteys. Kromosomihäiriöitä esiintyy 7:llä tuhannesta vastasyntyneestä, ja ne johtavat myös alkion kuolemaan (keskenmenoon) raskauden ensimmäisellä kolmanneksella puolessa tapauksista. Jos lapsi, jolla on kromosomipoikkeavuuksia, syntyy elävänä, kärsii yleensä vakavista vaivoista, jää jälkeen henkisessä ja fyysisessä kehityksessä.
Biokemialliset menetelmät . Sisällön avulla voit tunnistaa monia perinnöllisiä ihmisen sairauksia, jotka liittyvät aineenvaihduntahäiriöihin. Hiilihydraattien, aminohappojen, lipidien ja muun tyyppisten aineenvaihdunnan poikkeavuuksia tiedetään. Joten esimerkiksi diabetes mellitus johtuu haiman normaalin toiminnan rikkomisesta - se ei vapauta vaadittua määrää hormoni-insuliinia verenkiertoon, minkä seurauksena verensokeritaso nousee. Tämä häiriö ei johdu yhdestä ainoasta vakavasta geneettisen tiedon virheestä, vaan kokonaisesta pienistä virheistä, jotka yhdessä johtavat tai altistavat sairauteen.
Somaattisten solujen geneettiset menetelmät - tutkii somaattisten solujen perinnöllisyyttä ja vaihtelua, ts. kehon solut, ei sukupuoli. Somaattisilla soluilla on koko joukko geneettistä tietoa, ja niiden avulla voidaan tutkia koko organismin geneettisiä ominaisuuksia. Ihmisen somaattiset solut saadaan geneettiseen tutkimukseen biopsiamateriaalista (kudosten tai elinten intravitaalileikkaus), kun pieni pala kudosta otetaan tutkimukseen.
Immunogeneettiset menetelmät . Immunogeneettiseen menetelmään kuuluu serologisia menetelmiä, immunoelektroforeesia jne., joilla tutkitaan kudosten veriryhmiä, proteiineja ja seerumientsyymejä. Sen avulla voit todeta immunologisen yhteensopimattomuuden, tunnistaa immuunipuutoksen, kaksosten mosaiikismin jne.
Molekyyligeneettiset menetelmät . Menetelmien monipuolisuus. Tärkeimpien metodologisten lähestymistapojen (DNA:n uutto, restriktio, elektroforeesi, blottaus, hybridisaatio) karakterisointi. Polymeraasiketjureaktio, sekvensointi. Molekyyligeneettisten menetelmien mahdollisuudet ja laajuus perinnöllisen patologian diagnosoinnissa.
Menetelmät geenisidoksen tutkimiseen . Menetelmän perusteet ja soveltamisen ehdot ihmisgenetiikassa ja lääketieteellisessä genetiikassa.
Perinnöllisten sairauksien biologinen mallinnus tutkii ihmisten sairauksia eläimillä, jotka voivat sairastua näihin tauteihin. Se perustuu Vavilovin lakiin homologisista perinnöllisen vaihtelun sarjasta, esimerkiksi sukupuoleen liittyvää hemofiliaa voidaan tutkia koirilla, epilepsiaa kaneilla, diabetes mellitusta, lihasdystrofiaa rotilla, huulten ja kitalaen sulkeutumattomuutta hiirillä.
Lääketieteellinen geneettinen neuvonta - Erikoissairaanhoito on yleisin perinnöllisten sairauksien ehkäisymuoto. Geneettinen neuvonta - koostuu henkilölle tiedottamisesta perinnöllisen sairauden riskistä, sen välittämisestä jälkeläisille sekä diagnostisista ja terapeuttisista toimista.
Vaihe 1 neuvonta - taudin diagnoosin selventäminen.
Vaihe 2 neuvonta - sairastuneen lapsen riskin määrittäminen.
Vaihe 3 neuvonta - geneetin tulee tehdä johtopäätös tutkittavien lasten sairausriskistä ja antaa heille asianmukaiset suositukset.
4 (viimeinen) vaihe neuvonta - oikea vastaus ja odotetun raskauden mahdolliset komplikaatiot tai lopputulos heille ymmärrettävällä kielellä.
Tehtävä lääketieteellinen genetiikka on perinnöllisten sairauksien tunnistaminen, tutkiminen, ehkäisy ja hoito sekä keinojen kehittäminen ympäristötekijöiden haitallisten vaikutusten ehkäisemiseksi ihmisen perinnöllisyyteen.Sairauksia, joilla ei ole mitään tekemistä perinnöllisyyden kanssa, ei käytännössä ole olemassa. Ehdollisesti perinnölliset sairaudet voidaan jakaa kolmeen suureen ryhmään: aineenvaihduntataudit, molekyylisairaudet, jotka yleensä johtuvat geenimutaatioista, ja kromosomitaudit.
Geenimutaatiot voidaan ilmaista joidenkin entsyymien aktiivisuuden lisääntymisenä tai laskuna niiden puuttumiseen asti. Fenotyyppisesti tällaiset mutaatiot esiintyvät perinnöllisinä aineenvaihduntasairauksina, jotka määräytyvät vastaavan biokemiallisen reaktion tuotteen puuttumisen tai ylimäärän perusteella. Geenimutaatiot luokitellaan fenotyyppisen ilmenemismuotonsa mukaan eli sairauksiksi, jotka liittyvät aminohappo-, hiilihydraatti-, lipidi-, mineraali- ja nukleiinihappoaineenvaihdunnan rikkomiseen.
Kromosomitaudit. Tämän tyyppinen perinnöllinen sairaus liittyy muutokseen kromosomien lukumäärässä tai rakenteessa. Kromosomipoikkeavuuksien esiintymistiheys vastasyntyneillä vaihtelee välillä 0,6-1 %, ja 8-12 viikon vaiheessa niitä on noin 3 %:lla alkioista. Spontaanien keskenmenon joukossa kromosomipoikkeavuuksien esiintymistiheys on noin 30%, ja alkuvaiheessa (enintään kaksi kuukautta) - 50% ja enemmän. Ihmisillä on kuvattu kaiken tyyppisiä kromosomi- ja genomimutaatioita, mukaan lukien aneuploidia, jota voi olla kahta tyyppiä -myosomia ja polysomia. Monosomia on erityisen raskasta
Shereshevskyn oireyhtymä - Turner (44 + X), ilmenee naisilla, joille ovat ominaisia patologiset muutokset ruumiinrakenteessa (lyhyt kasvu, lyhyt kaula), lisääntymisjärjestelmän kehityksen häiriöt (useimpien naisten toissijaisten seksuaalisten ominaisuuksien puuttuminen), henkinen rajoittuneisuus. Tämän poikkeaman esiintymistiheys on 1: 4000-5000.
Trisomiaiset naiset (44 + XXX), yleensä ne erottuvat seksuaalisen, fyysisen ja henkisen kehityksen rikkomuksista, vaikka joillakin potilailla nämä merkit eivät välttämättä näy. On tunnettuja tapauksia tällaisten naisten hedelmällisyydestä. Oireyhtymän esiintymistiheys on 1:1000.
Klinefelterin oireyhtymä (44 + XXY) jolle on tunnusomaista sukurauhasten kehitys ja toiminta heikentynyt, eu-nuchoid -vartalotyyppi (kapeampi kuin lantio, hartiat, vartalon karvat ja rasvakerrostuminen vartalolle naistyypin mukaan, pitkulainen verrattuna käsivarsien ja jalkojen runkoon). Siksi korkeampi kasvu. Nämä merkit yhdistettynä jonkinlaiseen kehitysvammaisuuteen ilmenevät suhteellisen normaalilla pojalla murrosiästä lähtien. Klinefelterin oireyhtymää havaitaan polysomian yhteydessä paitsi X-kromosomissa (XXX XXXY, XXXXY), mutta myös Y-kromosomissa (XYY. XXYY. XXYYY). Oireyhtymän esiintymistiheys on 1:1000.
Downin oireyhtymä ( trisomia kromosomissa 21) . Eri kirjoittajien mukaan Downin syndroomaa sairastavien lasten syntymätaajuus on 1:500-700 vastasyntynyttä, ja viime vuosikymmeninä trisomia-21:n esiintymistiheys on lisääntynyt.
Sairaan lapsen syntyessä hänelle on joskus mahdollista saada lääkitystä, ruokavalio- ja hormonihoitoa. Poliomyeliitti voi toimia havainnollistavana esimerkkinä, joka vahvistaa lääketieteen mahdollisuudet torjua perinnöllisiä sairauksia. Tälle taudille on ominaista perinnöllinen taipumus, mutta taudin suora syy on virusinfektio. Massaimmunisointi taudin aiheuttajaa vastaan mahdollisti kaikkien perinnöllisesti altistuvien lasten pelastamisen taudin vakavilta seurauksilta. Ruokavalio- ja hormonihoitoa on käytetty menestyksekkäästi fenyyliketonurian, diabetes mellituksen ja muiden sairauksien hoidossa.
Ontogeneesi perinnöllisen tiedon toteutumisena tietyissä ympäristöolosuhteissa. Ontogeneesin päävaiheet. Ontogeneettisen kehityksen tyypit. Ontogeneesin periodisointi.
Ontogeneesi, tai kehon yksilöllistä kehitystä , suoritetaan hedelmöitykseen tulleiden vanhempien sukusolujen kautta saadun perinnöllisen ohjelman perusteella (suvuttoman lisääntymisen aikana tämä ohjelma on suljettu ainoan jälkeläisiä antavan vanhemman erikoistumattomiin soluihin). Perinnöllisen tiedon toteutumisen aikana ontogeneesiprosessissa organismi muodostaa lajin ja yksilölliset morfologiset, fysiologiset ja biokemialliset ominaisuudet, toisin sanoen - fenotyyppi. Kehitysprosessissa keho muuttaa luonnollisesti ominaisuuksiaan, mutta pysyy silti integroitu järjestelmä... Siksi fenotyyppi tulee ymmärtää ominaisuuksien kokonaisuutena koko yksilön kehityksen aikana, jonka jokaisessa vaiheessa on erityispiirteitä.
Johtava rooli fenotyypin muodostumisessa kuuluu perinnöllinen tieto, organismin genotyypin sisällä. Tässä tapauksessa yksinkertaiset ominaisuudet kehittyvät vastaavien alleelisten geenien tietyntyyppisen vuorovaikutuksen seurauksena. Samalla koko genotyyppijärjestelmällä on merkittävä vaikutus niiden muodostumiseen. Monimutkaisten ominaisuuksien muodostuminen tapahtuu ei-alleelisten geenien erilaisten vuorovaikutusten seurauksena suoraan genotyypissä tai niiden hallitsemissa tuotteissa. Tsygootin yksilöllisen kehityksen aloitusohjelma sisältää myös ns. spatiaaliset tiedot, jotka määrittelevät anteroposterioriset ja dorsaali-vatsa- (dorsoventraaliset) koordinaatit rakenteiden kehittymistä varten.
Tämän lisäksi yksilön genotyyppiin sisältyvän perinnöllisen ohjelman täytäntöönpanon tulos riippuu suurelta osin olosuhteista, joissa tämä prosessi suoritetaan. Ympäristön genotyypin ulkopuoliset tekijät voivat edistää tai estää geneettisen tiedon fenotyyppistä ilmenemistä, tehostaa tai heikentää sellaisen ilmenemisen astetta. Jo transkriptiovaiheessa yksittäisten geenien ilmentymistä ohjaa geneettisten ja ei-geneettisten tekijöiden vuorovaikutus. Näin ollen jopa organismin perusominaisuuksien - polypeptidien - muodostumisessa genotyyppi on mukana vuorovaikutteisten geenien järjestelmänä ja ympäristönä, jossa se toteutuu.
Yksilöllisen kehityksen genetiikassa keskiviikko on monimutkainen käsite. Yhtäältä tämä on välitön ympäristö, jossa yksittäiset geenit ja genotyyppi kokonaisuudessaan suorittavat tehtävänsä. Sen muodostavat kaikki kehon sisäisen ympäristön tekijöiden joukko: solusisältö (pois lukien DNA), suorien solujen välisten vuorovaikutusten luonne, biologisesti aktiiviset aineet (hormonit). Perinnöllisen ohjelman toteuttamiseen vaikuttavien organismien sisäisten tekijöiden joukko on nimetty Keskiviikkona 1. tilaus. Tämän ympäristön tekijät vaikuttavat erityisen voimakkaasti genotyypin toimintaan aktiivisten morfogeneettisten prosessien aikana, ensisijaisesti alkion synnyssä. Toisaalta ympäristön käsite erottuu, tai 2. tilauksen ympäristöt, kehon ulkopuolisten tekijöiden joukkona.
Ontogeneesin periodisointi Yksilöllinen kehitys on kokonaisvaltainen jatkuva prosessi, jossa yksittäiset tapahtumat linkitetään tilassa ja ajassa. Ontogenian periodisointiin on olemassa useita järjestelmiä, joista jokainen on sopivin tiettyjen tieteellisten tai käytännön ongelmien ratkaisemiseen.
KANSSA yleinen biologinen näkökulmat: lisääntymistä edeltävä, lisääntyväja nakuutti lisääntymiskyky.
V lisääntymistä edeltävä ajanjaksoa yksilö ei ole lisääntymiskykyinen. Sen pääsisältö on sukukypsän fenotyypin kehittäminen.
Alkionaikainen tai alkion, ontogeneesijakso alkaa hedelmöityksestä ja jatkuu, kunnes alkio poistuu munakalvoista.
Toukka ajanjaksoa havaitaan tyypillisesti niiden selkärankaisten kehityksessä, joiden alkiot nousevat ulos munakalvoista ja alkavat elää itsenäistä elämäntapaa saavuttamatta organisaation lopullisia (kypsiä) piirteitä.
metamorfoosi koostuu toukan muuttamisesta nuoruusmuotoon.
Nuorten ajanjakso alkaa muodonmuutoksen päättymishetkestä ja päättyy murrosikään ja lisääntymisen alkamiseen.
V lisääntymiskykyinen ajanjaksoa yksilö suorittaa seksuaalisen lisääntymisen tehtävän.
Jälkikasvu ajanjaksoa liittyy kehon ikääntymiseen ja sille on ominaista lisääntymiseen osallistumisen heikkeneminen tai täydellinen lopettaminen.
Ihmisen ontogeneettisyys
Antenataalinen ontogeneesi:
Idu- tai alkiokausi. Ensimmäinen viikko raskauden jälkeen.
Alkion aikakausi. Toinen on viides raskausviikko.
Sikiöjakso 32 viikkoa.
Synnytyksen jälkeinen ontogeneesi:
Vastasyntynyt tai vastasyntynyt ajanjakso. 1-10 päivää.
Rintojen ikä. 10 päivää - 1 vuosi.
Varhaislapsuus. 1-3 vuotta.
Ensimmäinen lapsuus. 4-7 vuotta vanha.
Toinen lapsuus. Pojille 8-12v, tytöille 8-11v.
Nuoruus. Pojille 13-16v, tytöille 12-15v.
Nuoruus. Pojille 17-21v, tytöille 16-20v.
Aikuinen ikä:
minäikä: 22-35-vuotiaat miehet, 21-35-vuotiaat naiset.
IIajanjakso: 36-60 vuotta miehellä, 36-55 vuotta naisella.
Vanhempi ikä. Miehet 61-74v, naiset 56-74v.
Seniili ikä. 75-90 vuotta vanha.
Pitkäikäisyysjakso. Yli 90 vuotta vanha.
Itämisaika on hetki hedelmöittymisen alusta alkion muodostumiseen. Alkiojakso on jaettu kahteen vaiheeseen: histotrofisen ravinnon vaiheeseen ja keltuaisen verenkierron vaiheeseen. Sikiön aikana tapahtuu siirtyminen keltuaisesta hemo-amniotrofiseen ravitsemukseen. Vastasyntyneen aikana vauva ruokkii ternimaitoa. Imetysaikana kypsät ja sen jälkeen täydentävät ruoat yhdistetään äidinmaitoon ja toteutetaan seisomisen sensorimotorinen järjestelmä. Kaudella varhaislapsuus siellä hallitaan kävely- ja puhetaidot. Ensimmäisessä lapsuudessa se kasvaa sanastoa ja ajattelun muodostumisen ensimmäinen vaihe etenee. Toisessa lapsuudessa aivojen analyyttinen ja synteettinen toiminta monimutkaistuu ja ajattelun toinen vaihe muodostuu. Teini-iässä sisäelinten kypsyminen on pääosin päättynyt ja ajattelun organisoitumisen kolmas vaihe etenee. Murrosikä tai murrosikä on käännekohta, jolloin persoonallisuuden muodostuminen ja murrosikä päättyvät. Kypsyyden tai vakauden aika on sosiaalisesti ja fysiologisten toimintojen organisoitumisen kannalta tuottavin. Vanhuuden aikana alkavat involuutiomuutokset, jotka ovat seurausta homeostaasin fysiologisista uudelleenjärjestelyistä.Seuraavina aikoina niistä tulee aktiivisia.
Ontogenian ja filogenian välinen korrelaatio. K. Beerin alkion samankaltaisuuden laki. E. Haeckelin ja F. Müllerin biogeneettinen laki
Ensimmäinen alkion samankaltaisuuden laki "Kun kuuluvien organismien varhaiset kehitysvaiheet eri luokat enemmän samankaltaisia kuin myöhemmät vaiheet."
2. laki kehityksen erikoistumisen "Ontogeneesin edetessä jokainen organismi kehittää yhä enemmän erityisiä ominaisuuksia."
F. Müller: "Evoluutiomuutokset rakenteessaaikuisiaeläimet johtuvatmuutoksia jälkeläisten ontogeneesin kulussaverrattuna heidän esi-isiensä ".
E. Haeckel Loi kolminkertaisen rinnakkaisuuden menetelmän:
vertaileva morfologia
vertailevat embryologiatiedot
paleontologiset tiedot
lähteitä fylogeneettisen sarjan rakentamiseen
Biogeneettinen laki"Ontogeny on nopea ja lyhyt fysiologian toisto"
Yhteenveto -se on toisto esi-isiensä evoluutiovaiheiden jälkeläisten ontogeniassa.
Ontogenian ja filogeneesin välinen suhde ... Nykyaikaisten käsitteiden mukaan useimmat fylogeneettiset innovaatiot liittyvät ontogeneettisiin heterokronioihin, eli erilaisten ontogeneettisten prosessien suhteellisten nopeuksien muutoksiin. Yksi evoluutionaalisesti merkittävimmistä heterokronioista on murrosiän siirtyminen evolutionaarisissa jälkeläisissä esi-isiensä toukkia vastaaviin vaiheisiin. Tällaista muutosta kutsutaan neoteniaksi tai pedomorfoosiksi. Tässä tapauksessa evolutionaaristen jälkeläisten elinkaari yleensä lyhenee (esimerkiksi esivanhemmille ominaisen metamorfoosivaiheen menettämisen vuoksi). Ei-sentimenttiä pidetään yhtenä tavoista saavuttaa nopeaa evoluution edistystä.
Ontogeneettisten ongelmien edelleen kehittäminen on äärimmäisen tärkeää sekä perusluonnontieteen että useiden lääketieteellisten, bioteknologisten ja ympäristöongelmien kannalta.
Alkion kehityksen päävaiheiden ominaisuudet ja merkitys: esitsygoottijakso, hedelmöitys, tsygootti, pilkkoutuminen. Niiden säätelymekanismit geneettisellä ja solutasolla.
Lannoitus - se on sukusolujen fuusioprosessi. Hedelmöityksen seurauksena muodostunut diploidisolu -tsygootti -edustaa Ensimmäinen taso uuden organismin kehittyminen. Lannoitusprosessi koostuu kolmesta peräkkäisestä vaiheesta:
a) sukusolujen lähentyminen (gamoneja(sukusolujen hormonit) toisaalta aktivoivat siittiöiden liikettä ja toisaalta niiden kiinnittymistä.akrosominen reaktio,jonka aikana munakalvot liukenevat akrosomin proteolyyttisten entsyymien vaikutuksesta. Lisäksi munasolun ja siittiön plasmakalvot sulautuvat yhteen ja molempien sukusolujen sytoplasman sytoplasman sillan kautta yhdistyvät. Sitten siittiön ydin ja sentrioli siirtyvät munan sytoplasmaan, ja siittiökalvo upotetaan munan kalvoon. Useimmissa eläimissä siittiön häntä pääsee myös munasoluun, mutta sitten se erottuu ja imeytyy ilman, että sillä on merkitystä jatkokehityksessä;
b) munan aktivointi Koska osa siittiöiden kalvosta läpäisee natriumioneja, viimeksi mainitut alkavat päästä munaan, mikä muuttaa solun kalvopotentiaalia. Sitten sukusolujen kosketuspisteestä etenevän aallon muodossa kalsiumionipitoisuuden kasvu tapahtuu, minkä jälkeen aivokuoren rakeet myös liukenevat aaltoon. Tässä tapauksessa vapautuvat spesifiset entsyymit edistävät vitelliinin kalvon irtoamista; se kovettuu, selannoituskuori.Kaikki kuvatut prosessit edustavat nsaivokuoren reaktio.;
c) sukusolujen fuusio eli syngamia Munasolu on siemennesteen kohtaamishetkellä yleensä jossakin meioosin vaiheista tietyn tekijän estämänä. Useimmissa selkärankaisissa tämä esto tapahtuu metafaasin II vaiheessa; monilla selkärangattomilla sekä kolmella nisäkäslajilla (hevonen, koira ja kettu) esto tapahtuu diakineesivaiheessa. Useimmissa tapauksissa meioosin esto vapautuu munasolun aktivoitumisen jälkeen hedelmöityksestä johtuen. Kun meioosi on valmis munassa, siihen tunkeutuneen siittiön ydin muuttuu. Se on interfaasin ja sitten profaasiytimen muodossa. Tänä aikana DNA kaksinkertaistuu jamiehen pronukleussaa vastaavan määrän perinnöllistä materiaaliaNS2 kanssa,nuo. sisältää haploidisen joukon replikoituneita kromosomeja. Munan ydin, joka on suorittanut meioosin, muuttuunaisen pronukleus,myös hankkimassaNS2 kanssa.Molemmat esiytimet tekevät monimutkaisia liikkeitä, sitten lähentyvät ja sulautuvat (synkario) , muodostaen yhteisen metafaasilevyn. Tämä on itse asiassa sukusolujen lopullisen fuusion hetki -syngamia.Tsygootin ensimmäinen mitoottinen jakautuminen johtaa kahden alkiosolun (blastomeerin) muodostumiseen, joissa on sarja kromosomeja 2n2 ckaikissa.
Tsygootti - diploidi(sisältää täydellisen kaksoissarjankromosomit) tuloksena muodostunut solulannoitus(fuusiotmunasolutjasiittiöitä). Tsygootti ontotipotentti(eli kykenee synnyttämään muita)häkki.
Ihmisellä on ensimmäinenmitoottinentsygootin jakautuminen tapahtuu noin 30 tuntia hedelmöityksen jälkeen, mikä johtuu monimutkaisista valmisteluprosesseista ensimmäiseen fragmentointiin. Tsygootin pilkkomisen seurauksena muodostuneita soluja kutsutaan
blastomeerit... Tsygootin ensimmäisiä jakautumisia kutsutaan "katkaisuiksi", koska solu jakautuu tarkasti: jokaisen jakautumisen jälkeen tytärsolut pienenevät ja jakautuvat, ja jakautumisten välillä ei ole solun kasvuvaihetta.
Erota - tämä on sarja tsygootin ja muiden blastomeerien peräkkäisiä mitoottisia jakautumisia, jotka päättyvät monisoluisen alkion muodostumiseen -blastula. Peräkkäisten jakautumisten välillä ei tapahdu solujen kasvua, mutta DNA syntetisoituu välttämättä. Kaikki DNA:n esiasteet ja välttämättömät entsyymit kertyvät oogeneesin aikana. Aluksi blastomeerit ovat vierekkäin muodostaen soluklusterin, jota kutsutaanmorula . Sitten solujen väliin muodostuu ontelo -blastocel, täynnä nestettä. Solut työnnetään reuna-alueille muodostaen blastula-seinän -blastodermi. Alkion kokonaiskoko katkaisun lopussa blastulavaiheessa ei ylitä tsygootin kokoa. Fragmentointijakson tärkein tulos on tsygootin muuttuminenmonisoluinen yksikerroksinen alkio .
Pilkkomisen morfologia. Yleensä blastomeerit sijaitsevat tiukassa järjestyksessä suhteessa toisiinsa ja munan napa-akseliin. Murskausjärjestys tai -menetelmä riippuu keltuaisen määrästä, tiheydestä ja jakautumisesta munassa. Sachs - Hertwigin sääntöjen mukaan soluytimellä on taipumus sijaita sytoplasman keskellä ilman keltuaista ja solunjakautumisen kara - tämän vyöhykkeen suurimman laajuuden suuntaan.
Oligo- ja mesolesiittinen munat murskaamallasaattaa loppuun,taiholoblastinen.Tämän tyyppinen pilkkoutuminen löytyy nahkiaisista, joistakin kaloista, kaikista sammakkoeläimistä sekä pussieläimistä ja istukan nisäkkäistä. Täydellisen murskaamisen yhteydessä ensimmäisen jaon taso vastaa kahdenvälisen symmetriatasoa. Toisen jaon taso kulkee kohtisuorassa ensimmäisen jaon tasoon nähden. Kahden ensimmäisen jaon molemmat uurteet ovat meridiaaneja, ts. alkaa eläinnapasta ja leviää kasvulliseen napaan. Munasolu on jaettu neljään blastomeeriin, jotka ovat suurin piirtein samankokoisia. Kolmannen jaon taso kulkee leveyssuunnassa kohtisuorassa kahteen ensimmäiseen nähden. Tämän jälkeen mesolecitaalisissa munissa esiintyy epätasaista pilkkoutumista kahdeksan blastomeerin vaiheessa. Eläinpaalussa on neljä pienempää blastomeeria -mikrometriä,vegetatiivisella - neljä isompaa -makromeerit.Sitten jako jatkuu jälleen meridiaanitasoilla ja sitten taas leveystasoilla.
Monikielisessä muodossa teleost-kalojen, matelijoiden, lintujen, sekä yksiväristen nisäkkäiden munasolut murskaavatosittainen,taimerob-elastinen,nuo. kattaa vain keltuaisen sytoplasman. Se sijaitsee ohuen kiekon muodossa eläinpaalussa, joten tämän tyyppistä murskaamista kutsutaankiekkomainen.Katkaisutyyppiä karakterisoitaessa otetaan huomioon myös blastomeerien suhteellinen sijainti ja jakautumisnopeus. Jos blastomeerit on järjestetty riveihin toistensa päälle säteitä pitkin, pilkkominen on ns.säteittäinen.Se on tyypillistä chordaateille ja piikkinahkaisille. Luonnossa on muitakin muunnelmia blastomeerien avaruudellisesta järjestelystä katkaisun aikana, mikä määrittää sen tyypit, kuten spiraali nilviäisissä, bilateraalinen sukkulamadoissa ja anarkkinen meduusoissa.
Keltuaisen jakautumisen ja eläin- ja kasviperäisten blastomeerien jakautumisen synkronisuusasteen välinen suhde havaittiin. Piikkinahkaisten oligolesitaalisissa munissa pilkkoutuminen on lähes synkronista; mesolesitaalisissa munasoluissa synkronisuus on heikentynyt kolmannen jakautumisen jälkeen, koska autonomiset blastomeerit johtuvat suuri numero keltuainen jakautuu hitaammin. Lomakkeissa, joissa on osittainen pirstoutuminen, jaot ovat asynkronisia alusta alkaen jakeskeisellä paikalla olevat blastomeerit jakautuvat nopeammin.
Murskauksen lopussa muodostuu blastula. Blastulan tyyppi riippuu pilkkoutumistyypistä ja siten munatyypistä.
Molekyyligeneettisten ja biokemiallisten prosessien ominaisuudet murskaamisen aikana. Kuten edellä todettiin, mitoottiset syklit katkaisujakson aikana lyhenevät suuresti, erityisesti aivan alussa.
Esimerkiksi merisiilin munissa koko fissiosykli kestää 30–40 minuuttia, kun S-vaihe kestää vain 15 minuuttia. GI- jaG2-jaksot ovat käytännössä poissa, koska munasolun sytoplasmaan on luotu tarvittava tarjonta kaikista aineista, ja mitä suurempi, sitä suurempi se on. Ennen jokaista jakautumista syntetisoidaan DNA ja histonit.
Replikaatiohaarukan etenemisnopeus DNA:ta pitkin katkaisun aikana on normaali. Samaan aikaan blastomeeri-DNA:ssa havaitaan enemmän aloituspisteitä kuin somaattisissa soluissa. DNA-synteesi tapahtuu kaikissa replikoneissa samanaikaisesti, synkronisesti. Siksi DNA:n replikaatioaika ytimessä osuu yhteen yhden, lisäksi lyhentyneen replikonin, kaksinkertaistumisajan kanssa. On osoitettu, että kun tuma poistetaan tsygootista, tapahtuu pilkkoutuminen ja alkio saavuttaa kehityksessään lähes blastula-vaiheen. Jatkokehitys pysähtyy.
Pilkkomisen alussa muun tyyppinen ydinaktiivisuus, esimerkiksi transkriptio, puuttuu käytännössä. Erilaisissa munatyypeissä geenin transkriptio ja RNA-synteesi alkavat eri vaiheissa. Tapauksissa, joissa sytoplasmassa on monia erilaisia aineita, kuten esimerkiksi sammakkoeläimissä, transkriptio ei aktivoidu heti. RNA-synteesi niissä alkaa varhaisessa blastula-vaiheessa. Sitä vastoin nisäkkäillä RNA-synteesi alkaa jo kahden blastomeerin vaiheessa.
Katkaisujakson aikana muodostuu RNA:ta ja proteiineja, jotka ovat samanlaisia kuin oogeneesiprosessissa syntetisoituneet. Nämä ovat pääasiassa histoneja, solukalvojen proteiineja ja solun jakautumiseen tarvittavia entsyymejä. Näitä proteiineja käytetään välittömästi yhdessä aiemmin munasolujen sytoplasmaan varastoitujen proteiinien kanssa. Tämän lisäksi katkaisujakson aikana on mahdollista synteesi proteiineja, joita ei ennen ollut olemassa. Tätä tukevat tiedot alueellisista eroista RNA:n ja proteiinien synteesissä blastomeerien välillä. Joskus nämä RNA:t ja proteiinit alkavat toimia myöhemmässä vaiheessa.
Tärkeä rooli pilkkomisessa on sytoplasman jakautumisella -sytotomia.Sillä on erityinen morfogeneettinen merkitys, koska se määrittää pilkkoutumistyypin. Sytotomian prosessissa muodostuu ensin supistuva mikrofilamenttirenkaan avulla. Tämän renkaan kokoaminen tapahtuu mitoottisen karan napojen suoran vaikutuksen alaisena. Sytotomian jälkeen oligolesitaalisten munien blastomeerit pysyvät yhdistettyinä vain ohuilla silloilla. Juuri tähän aikaan ne on helpoin erottaa toisistaan. Tämä johtuu siitä, että sytotomia johtaa solujen välisen kosketuspinta-alan pienenemiseen rajallisen kalvopinta-alan vuoksi, heti sytotomian jälkeen alkaa uusien solun pinnan osien synteesi, kosketuspinta-ala kasvaa ja blastomeerit alkavat koskettaa tiukasti. Halkeamisuurteet kulkevat erillisten ovoplasman alueiden välisiä rajoja pitkin, mikä heijastaa munasolujen segregaation ilmiötä.Siksi eri blastomeerien sytoplasman kemiallinen koostumus eroaa.
Alkion kehityksen päävaiheiden ominaisuudet ja merkitys: gastrulaatio, histo- ja organogeneesi. 2- ja 3-kerroksisten alkioiden muodostuminen. Menetelmät mesodermin muodostumiseen. Alkiokerrosten johdannaiset. Näiden prosessien säätelymekanismit geneettisellä ja solutasolla.
Histogeneesi - (kreikan sanasta histos - kudos it ... genesis), filogeniassa kehittynyt prosessisarja, joka tarjoaa monisoluisten organismien ontogeneesissä kudosten muodostumisen, olemassaolon ja palautumisen niiden luontaisesti elinkohtaisina. ominaisuudet. Kehossa kudokset kehittyvät määritelmän mukaan. alkion alkuvaiheet (alkiokerrosten johdannaiset), jotka muodostuvat alkiosolujen lisääntymisen, liikkeen (morfogeneettiset liikkeet) ja adheesion seurauksena sen kehityksen varhaisessa vaiheessa organogeneesiprosessissa. Olennot, tekijä G. - määrättyjen solujen erilaistuminen, mikä johtaa useiden morfolin esiintymiseen. ja fiziol. solutyyppejä, jotka jakautuvat säännöllisesti kehossa. Joskus G.:een liittyy solujen välisen aineen muodostuminen. Tärkeä rooli G.:n suunnan määrittämisessä kuuluu solujen välisille kontaktivuorovaikutuksille ja hormonaalisille vaikutuksille. Joukko soluja, jotka tekevät määrityksiä. G., on jaettu useisiin ryhmiin: esi-isien (kanta)solut, jotka pystyvät erilaistumaan ja täydentämään oman lajinsa menetystä jakautumalla; progenitorisolut (ns. puolikantasolut) - erilaistuvat, mutta säilyttävät kyvyn jakautua; kypsä differenti. soluja. Reparative G. synnytyksen jälkeisessä jaksossa on vaurioituneiden tai osittain kadonneiden kudosten palauttamisen taustalla. Ominaisuudet, G.:n muutokset voivat johtaa kasvaimen syntymiseen ja kasvuun.
Organogeneesi (kreikasta.organoni- urut,synty- kehitys, koulutus) - elinten kehitys- tai muodostumisprosessi ihmisten ja eläinten alkiossa. Organogeneesi seuraa aikaisempia alkion kehityksen jaksoja (katso alkio) - munasolujen murskaamista, gastrulaatiota ja tapahtuu sen jälkeen, kun elinten ja kudosten tärkeimmät alkiot (anlages) on eristetty. Organogeneesi etenee rinnakkain histogeneesin (katso) tai kudoskehityksen kanssa. Toisin kuin kudoksissa, joista jokainen on peräisin jostakin alkion alkuaineesta, elimet syntyvät yleensä useiden (kahdesta neljään) erilaisen esikuoren osallistuessa (katso itukerrokset), jolloin syntyy erilaisia kudoskomponentteja. urut. Esimerkiksi suolen seinämän osana elimen onkaloa vuoraava epiteeli ja rauhaset kehittyvät sisäisestä itukerroksesta - endodermista (katso), sidekudoksesta verisuonilla ja sileästä lihaskudoksesta - mesenkyymistä (katso), suolen seroosikalvoa peittävä mesoteeli - splanknotomin viskeraalisesta levystä eli keskimmäisestä itukerroksesta - mesodermista ja elimen hermoista ja ganglioista - hermokipusta. Iho muodostuu ulkoisen itukerroksen - ektodermin (katso), josta orvaskesi ja sen johdannaiset (hiukset, tali- ja hikirauhaset, kynnet jne.) kehittyvät, sekä dermatomit, joista mesenkyymi syntyy, erilaistuen. ihon sidekudospohjaan (dermis). Hermot ja hermopäätteet ihossa, kuten muuallakin, ovat hermoprimordiumin johdannaisia. Jotkut elimet muodostuvat yhdestä primordiumista, esimerkiksi luu, verisuonet, imusolmukkeet - mesenkyymistä; kuitenkin myös täällä alkuhermoston johdannaiset - hermosäikeet - kasvavat anlageen ja muodostuu hermopäätteitä.
Jos histogeneesi koostuu pääasiassa solujen lisääntymisestä ja erikoistumisesta sekä solujenvälisten aineiden ja muiden ei-solurakenteiden muodostumisesta niiden avulla, niin organogeneesin taustalla olevat pääprosessit ovat poimujen, ulkonemien, ulkonemien, paksuuntumisen, epätasaisen kasvun muodostuminen. , itukerrosten lisääntyminen tai erottuminen (eristys), sekä erilaisten kirjanmerkkien keskinäinen itäminen. Ihmisillä organogeneesi alkaa kolmannen viikon lopussa ja päättyy yleisesti kohdunsisäisen kehityksen 4. kuukauteen mennessä. Useiden alkion väliaikaisten (väliaikaisten) elinten - korioni, amnion, keltuaispussi - kehitys alkaa kuitenkin jo 1. viikon lopulla, ja jotkut lopulliset (lopulliset) elimet muodostuvat myöhemmin kuin toiset (esim. imusolmukkeet - kohdunsisäisen kehityksen viimeisistä kuukausista murrosiän alkamiseen).
Gastrulaatio - yksikerroksinen alkio - blastula - muuttuumonikerroksinen -kaksi- tai kolmikerroksinen, nsgastrula(kreikasta.gasteri -vatsa pienenevässä merkityksessä).
Alkuperäisissä chordaateissa, esimerkiksi lansetissa, homogeeninen yksikerroksinen blastodermi muuttuu gastrulaation aikana ulommaksi itukerrokseksi - ektodermiksi - ja sisäiseksi itukerrokseksi -endodermi.Endodermi muodostaa ensisijaisen suolen, jonka sisällä on onkalogastrocoel.Gastrocoeliin johtavaa reikää kutsutaanblastoporetai ensisijainen suu.Kaksi alkiokerrostaovat gastrulaation määritteleviä morfologisia merkkejä. Niiden olemassaolo tietyssä kehitysvaiheessa kaikissa monisoluisissa eläimissä, coelenterateista korkeampiin selkärankaisiin, mahdollistaa pohtimisen itukerrosten homologiasta ja kaikkien näiden eläinten alkuperän yhtenäisyydestä. Selkärankaisilla muodostuu gastrulaation aikana mainittujen kahden lisäksi kolmas alkiokerros -mesoderma,miehittää paikan ekto- ja endodermin välissä. Keskimmäisen alkion kerroksen, joka on chordomesodermi, kehitys on selkärankaisten gastrulaatiovaiheen evoluution mukainen komplikaatio, ja se liittyy niiden kehityksen kiihtymiseen alkion alkuvaiheissa. Primitiivisemmissä sointuissa, kuten lansetissa, chordomesoderma muodostuu yleensä gastrulaation jälkeisen vaiheen alussa -organogeneesi.Jälkeläisten joidenkin elinten kehitysajan muutos suhteessa muihin verrattuna esi-isien ryhmiin on ilmentymäheterokronia.Muutokset evoluutioprosessin tärkeimpien elinten munimisajassa eivät ole harvinaisia.
Gastrulaatioprosessille on ominaistatärkeitä solumuutoksia,kuten ryhmien ja yksittäisten solujen suunnattu liike, solujen selektiivinen lisääntyminen ja lajittelu, sytodifferentioitumisen ja induktiovuorovaikutusten alkaminen.
Gastrulaatiomenetelmät ovat erilaisia. Avaruuteen suunnattuja soluliikkeitä on neljää tyyppiä, mikä johtaa alkion muuttumiseen yksikerroksisesta monikerroksiseksi.
Intussusceptio - blastodermin yhden osan invaginaatio kokonaisen kerroksen sisällä. Lansetissa vegetatiivisen navan solut tunkeutuvat, sammakkoeläimissä invaginaatio tapahtuu eläimen ja kasvullisen navan välisellä rajalla harmaan sirpin alueella. Intussusseptioprosessi on mahdollista vain munissa, joissa on pieni tai keskimääräinen keltuainen.
Epibolia - eläinnavan pienten solujen liikakasvu, suurempien, jakautumisnopeudessa jäljessä ja vegetatiivisen navan vähemmän liikkuvia soluja. Tämä prosessi on selvä sammakkoeläimissä.
Nimitys - blastodermisolujen kerrostuminen kahdeksi kerrokseksi, jotka sijaitsevat toistensa päällä. Delaminaatiota voidaan havaita alkioiden diskoblastulassa, jossa on osittainen pilkkoutuminen, kuten matelijat, linnut ja munasolut nisäkkäät. Delaminaatio ilmenee istukan nisäkkäiden alkioblasteissa, mikä johtaa hypoblastin ja epiblastin muodostumiseen.
Maahanmuutto - ryhmien tai yksittäisten solujen liikkuminen, joita ei ole yhdistetty yhdeksi kerrokseksi. Maahanmuuttoa tapahtuu kaikissa alkioissa, mutta se on tyypillisintä korkeampien selkärankaisten gastrulaation toiselle vaiheelle. Jokaisessa erityisessä alkion synnyn tapauksessa yhdistetään yleensä useita gastrulaatiomenetelmiä.
Gastrulaation morfologia. Blastulan alueelle, jonka solumateriaalista gastrulaation ja varhaisen organogeneesin (neurulaation) aikana muodostuu yleensä täysin selvät itukerrokset ja -elimet. Intussusseptio alkaa vegetatiivisesta napasta. Nopeamman jakautumisen ansiosta eläinnavan solut kasvavat ja työntävät vegetatiivisen navan solut blastulaan. Tätä helpottaa sytoplasman tilan muutos soluissa, jotka muodostavat blastoporen huulet ja niiden vieressä. Intussusseptiosta johtuen blastokoeli pienenee ja gastrocoel lisääntyy. Samanaikaisesti blastokoelin katoamisen kanssa ektodermi ja endodermi joutuvat läheiseen kosketukseen. Lanseletissa, kuten kaikissa deuterostomeissa (niihin kuuluvat piikkinahkainen tyyppi, Chordate-tyyppi ja jotkut muut pienet eläintyypit), blastohuokosalue muuttuu kehon häntäosaksi, toisin kuin protostomeissa, joissa blastohuoko vastaa pään osaan. Deuterostomien suuaukko muodostuu alkion päähän vastapäätä blastohuokosta. Sammakkoeläinten gastrulaatiolla on paljon yhteistä lanseletin gastrulaation kanssa, mutta koska niiden munasoluissa on paljon enemmän keltuaista ja se sijaitsee pääasiassa vegetatiivisessa navassa, suuret amfiblastulan blastomeerit eivät pysty tunkeutumaan sisäänpäin.Intussusceptio menee vähän toisin. Eläimen ja kasvullisen napojen rajalla harmaan sirpin alueella solut venyvät ensin voimakkaasti sisäänpäin ottamalla muotoa≪ pullon muotoinen≫ ja vedä sitten blastulan pintakerroksen solut mukanaan. Näkyviin tulee sirpin muotoinen ura ja blastohuokosen selkähuuli. Samanaikaisesti eläinnavan pienemmät solut, jotka jakautuvat nopeammin, alkavat siirtyä kohti kasvullista napaa. Selkähuulen alueella ne kiertyvät ja työntyvät esiin, ja sivuilta ja puolikuun uraa vastakkaiselta puolelta kasvaa suurempia soluja. Sitten prosessiepiboli johtaa blastoporen sivu- ja vatsahuulten muodostumiseen. Blastohuoko sulkeutuu renkaaksi, jonka sisällä on jonkin aikaa näkyvissä vegetatiivisen navan suuret valosolut niin sanotun keltuaisen tulpan muodossa. Myöhemmin ne uppoavat kokonaan sisäänpäin ja blastohuokos kapenee. Sammakkoeläinten elintärkeillä (vitaalisilla) väriaineilla leimausmenetelmällä tutkittiin yksityiskohtaisesti blastulasolujen liikkeitä gastrulaation aikana.oletettu(Lat. praesumptio - olettamus), normaalin kehityksen aikana ne sisältyvät ensin tiettyihin elinten alkuaineisiin ja sitten itse elimiin. Tiedetään, että hännänttömillä sammakkoeläimillä oletetun jänteen ja mesoderman materiaali blastulavaiheessa ei ole sen pinnalla, vaan amfiblastulan seinämän sisäkerroksissa, kuitenkin suunnilleen samalla tasolla kuin kuvassa. Sammakkoeläinten varhaisten kehitysvaiheiden analyysi antaa meille mahdollisuuden päätellä tämänovoplasminen segregaatio,Se, joka ilmenee selvästi munasolussa ja tsygootissa, on erittäin tärkeä määritettäessä solujen kohtaloa, jotka ovat perineet tämän tai toisen sytoplasman osan. Gastrulaatiolla alkioissa, joissa on meroblastinen pilkkoutuminen ja kehitys, on omat ominaisuutensa. Omistaalintujase alkaa blastulan katkeamisen ja muodostumisen jälkeen alkion kulkiessa munanjohtimen läpi. Kun muna munitaan, alkio koostuu jo useista kerroksista: yläkerros on nsepiblastooma,pohja -primaarinen hypoblastooma.Niiden välillä on kapea rako - blastocoel. Sitten muodostettiinsekundaarinen hypoblasti,joiden muodostumistapa ei ole täysin selvä. On näyttöä siitä, että primaariset sukusolut ovat peräisin lintujen primaarisesta hypoblastista ja sekundaarinen muodostaa alkion ulkopuolisen endodermin. Primaaristen ja sekundaaristen hypoblastien muodostumista pidetään gastrulaatiota edeltävänä ilmiönä. Tärkeimmät gastrulaatiotapahtumat ja kolmen itukerroksen lopullinen muodostuminen alkavat munanpoiston jälkeen inkuboinnin alkaessa. Solujen kerääntyminen tapahtuu epiblastin takaosaan, mikä johtuu solujen epätasaisesta jakautumisnopeudesta ja niiden liikkumisesta epiblastin lateraalisista osista keskustaan, toisiaan kohti. Niin kutsuttuensisijainen nauha,joka ulottuu päätä kohti. Ensisijaisen nauhan keskelle muodostuuensisijainen ura,ja reunoja pitkin - ensisijaiset rullat. Ensisijaisen nauhan päädyssä tapahtuu paksuuntumista -Hensenin solmu,ja siinä on ensisijainen kuoppa. Kun epiblastisolut tulevat ensisijaiseen uraan, niiden muoto muuttuu. Ne muistuttavat muodoltaan≪ pullon muotoinen≫ sammakkoeläinten gastrulasolut. Nämä solut ottavat sitten tähtimuodon ja uppoavat epiblastin alle muodostaen mesodermin. Endodermi muodostuu primaaristen ja sekundaaristen hypoblastien pohjalta, ja siihen on lisätty uuden sukupolven endodermaalisia soluja, jotka siirtyvät blastodermin ylemmistä kerroksista. Useiden sukupolvien endodermaaliset solut osoittavat gastrulaatiojakson pidentymistä ajan myötä. Osa epiblastista Hensenin kyhmyn kautta kulkevista soluista muodostaa tulevan notochordin. Samanaikaisesti notokordin alkamisen ja pidentymisen kanssa Hensen-solmu ja ensisijainen raita häviävät vähitellen pään ja kaudaalisen pään suunnassa. Tämä vastaa blastohuokosen kapenemista ja sulkeutumista. Kun se supistuu, ensisijainen raita jättää taakseen alkion aksiaalisten elinten muodostuneet alueet päästä hännän suuntaan. Vaikuttaa järkevältä pitää kananpoikasalkion soluliikkeitä homologisina epiboleina ja ensisijaista raitaa ja Hensenin kyhmyä homologisina sammakkoeläinten gastrulan selkähuulen blastohuokosen kanssa. On mielenkiintoista huomata, että nisäkäsalkioiden solut säilyttävät gastrulaatiovaiheessa matelijoiden ja lintujen alkioiden tyypilliset liikkeet huolimatta siitä, että näissä eläimissä munissa on vähän keltuaista ja pilkkoutuminen on täydellinen. . Tämä vahvistaa ajatuksen nisäkkäiden alkuperästä esi-isien ryhmästä, jossa munat olivat runsaasti keltuaista.
Gastrulaatiovaiheen ominaisuudet. Gastrulaatiolle on ominaista erilaiset soluprosessit. Mitoosi jatkuusolujen lisääntyminen,lisäksi sillä on erilainen voimakkuus alkion eri osissa. Kuitenkin eniten ominaisuus gastrulaatio koostuusolumassojen liikkuminen.Tämä johtaa muutokseen alkion rakenteessa ja sen muuttumiseen blastulasta gastrulaksi. On tapahtumassalajittelusoluja kuulumalla eri itukerroksiin, joiden sisällä ne≪ tunnistaa≫ toisiaan. Gastrulaatiovaihe alkaasytodifferentiaatio,mikä tarkoittaa siirtymistä oman genomin biologisen tiedon aktiiviseen käyttöön. Yksi geneettisen aktiivisuuden säätelijöistä on alkiosolujen sytoplasman erilainen kemiallinen koostumus, joka muodostuu ovoplasmisen segregaation seurauksena. Sammakkoeläinten ektodermaaliset solut ovat siis tummia munan eläinnapasta niihin päätyneen pigmentin vuoksi, ja endodermin solut ovat kevyitä, koska ne ovat peräisin munan vegetatiivisesta napasta. Gastrulaation aikana roolialkion induktio.On osoitettu, että primaarisen juovan ilmaantuminen linnuissa johtuu hypoblastin ja epiblastin välisestä induktiivisesta vuorovaikutuksesta. Hypoblastille on ominaista polariteetti. Hypoblastin sijainnin muutos suhteessa epiblastiin aiheuttaa muutoksen primääriliuskan orientaatiossa. Kaikki luetellut prosessit kuvataan yksityiskohtaisesti luvussa. On huomattava, että tällaiset ilmentymäteheysalkion kaltainenmäärittely, alkion säätelyjaliittäminenovat hänelle luontaisia gastrulaation aikana samassa määrin kuin pilkkomisen aikana.
Mesoderman muodostuminen -Kaikilla eläimillä, paitsi coelenteraatilla, gastrulaation yhteydessä (samanaikaisesti sen kanssa tai seuraavassa vaiheessa gastrulaation vuoksi) ja kolmas alkiokerros - mesoderma. Tämä on kokoelma soluelementtejä, jotka sijaitsevat ektodermin ja endodermin välissä, eli blastoseleissa. Tällä tavalla. Siten alkiosta ei tule kaksikerroksista, vaan kolmikerroksista. Korkeammissa selkärankaisissa alkioiden kolmikerroksinen rakenne syntyy jo gastrulaation prosessissa, kun taas alemmissa sointuissa ja kaikissa muissa tyypeissä itse gastrulaation seurauksena muodostuu kaksikerroksinen alkio.
On mahdollista luoda kaksi pohjimmiltaan erilaista reittiä mesoderman syntymiselle: teloblastinen, erikoinen Protostomia, ja enterokoelous, ominaistaDeute-rosiomia. gastrulaation aikana protostomissa, ektodermin ja endodermin välisellä rajalla, blastohuokosen sivuilla, on jo kaksi suurta solua, jotka erottuvat itsestään (jakautumisesta johtuen) pieniä soluja. Siten muodostuu keskikerros - mesoderma. Teloblastit, jotka tuottavat uusia mesodermisolujen sukupolvia, jäävät alkion takapäähän. Tästä syystä tätä mesoderman muodostusmenetelmää kutsutaan teloblastinen (kreikan sanasta telos - loppu).
Enterocoele-menetelmällä muodostuvan mesodermin solujoukko esiintyy primäärisuolen taskumaisten ulkonemien muodossa (sen seinämien ulkoneminen blastokohteeseen). Nämä ulkonemat, joiden sisään ensisijaisen suolen ontelon alueet tulevat, erotetaan suolesta ja erotetaan siitä pussien muodossa. Pussien ontelo muuttuu yleensä ottaen, eli toissijaisessa ruumiinontelossa coelomic pussit voidaan jakaa keskimmäisen alkion kerroksen segmentteihin ei heijasta koko vaihteluiden ja poikkeamien vaihtelua, joka on täysin luonnollista yksittäisille eläinryhmille. Samoin kuin teloblastinen, mutta vain ulkoisesti, mesoderman muodostusmenetelmä ei ole teloblastien jakautuminen, vaan parittoman tiheän primordiumin (soluryhmän) ilmaantuminen blastohuokosen reunoihin, joka jakautuu myöhemmin kahdeksi symmetriseksi nauhaksi. soluja. Enterocoelous-menetelmällä mesoderman alkuosa voidaan yhdistää tai parittamaton; joissakin tapauksissa muodostuu kaksi symmetristä coelomic pussia, ja toisissa yksi yhteinen coelomic pussi muodostuu ensin, joka jakautuu myöhemmin kahdeksi symmetriseksi puolikkaaksi.
Alkiokerrosten johdannaiset. Kolmen itukerroksen tuleva kohtalo on erilainen.
Ektodermista kehittyy: kaikki hermokudos; ihon ulkokerrokset ja sen johdannaiset (hiukset, kynnet, hammaskiille) ja osittain suuontelon limakalvot, nenäontelot ja peräaukko.
Endodermista syntyy koko ruoansulatuskanavan - suuontelosta peräaukon - limakalvo ja kaikki sen johdannaiset, ts. kateenkorva, kilpirauhanen, lisäkilpirauhanen, henkitorvi, keuhkot, maksa ja haima.
Mesodermista muodostuu: kaikki tyypit sidekudos, luu- ja rustokudos, veri ja verisuonijärjestelmä; kaikki tyypit lihaskudos; eritys- ja lisääntymisjärjestelmät, ihon dermaalinen kerros.
Aikuisella eläimellä on hyvin vähän sellaisia endodermaalista alkuperää olevia elimiä, jotka eivät sisällä ektodermista peräisin olevia hermosoluja. Jokainen tärkeä elin sisältää myös mesodermin johdannaisia - verisuonia, verta ja usein lihaksia, joten itukerrosten rakenteellinen eristys säilyy vain niiden muodostumisvaiheessa. Jo kehityksensä alussa kaikki elimet saavat monimutkaisen rakenteen, ja ne sisältävät kaikkien itukerrosten johdannaisia
Ontogeneesin postembryonaalinen ajanjakso. Pääprosessit: kasvu, lopullisten rakenteiden muodostuminen, murrosikä, lisääntyminen, ikääntyminen.
Synnytyksen jälkeinen ontogeneesi - organismin kehitysvaihe syntymähetkestä kuolemaan. Siinä yhdistyvät kaksi vaihetta: a) varhaisen postnataalisen ontogeneesin vaihe; b) myöhäisen postnataalisen ontogeneesin vaihe. Varhainen postnataalinen ontogeneesi alkaa organismin syntymästä ja päättyy kaikkien elinjärjestelmien rakenteellisen ja toiminnallisen kypsymisen alkamiseen, mukaan lukien lisääntymisjärjestelmä... Sen kesto ihmisillä on 13-16 vuotta. Varhainen postnataalinen ontogeneesi voi sisältää organogeneesin, erilaistumisen ja kasvun pääprosessit (esimerkiksi kenguruilla) tai vain myöhemmin kypsyvien elinten (sukurauhaset, toissijaiset sukupuoliominaisuudet) kasvun ja erilaistumisen. Monilla eläimillä metamorfoosi tapahtuu alkion jälkeisessä kehityksessä. Myöhäinen postnataalinen ontogeneettisyys sisältää aikuisuuden, ikääntymisen ja kuoleman. Postembryoniselle kehitykselle on tunnusomaista: 1) intensiivinen kasvu; 2) lopullisten (lopullisten) ruumiinsuhteiden vahvistaminen; 3) elinjärjestelmien asteittainen siirtyminen kypsälle organismille ominaiseen toimintaan.
Korkeus - tämä on yksilön (organismin) massan ja lineaaristen mittojen kasvu, joka johtuu massan lisääntymisestä, mutta pääasiassa solujen lukumäärästä sekä ei-solumuodostelmista. Kasvukäyriä käytetään kuvaamaan kasvua (painon tai pituuden muutos ontogeneesin aikana), absoluuttisen ja suhteellisen kasvun indikaattoreita tietyn ajanjakson aikana sekä spesifistä kasvunopeutta.
Yksilön kasvulle on ominaista jompikumpiisometrinen - kehon osien ja elinten tasainen kasvu taiallometria - kehon osien epätasainen kasvu.Allometria Se voi olla negatiivinen (esimerkiksi lapsen pään hitaampi kasvu suhteessa vartaloon) ja positiivinen (esimerkiksi märehtijöiden sarvien kiihtynyt kasvu). Kasvuvauhti hidastuu iän myötä. Epämääräisen kasvun eläimet kasvavat koko elämänsä ajan (nilviäiset, äyriäiset, kalat, sammakkoeläimet). Eläimillä, joilla on tietty korkeus, tiettyyn ikään mennessä kasvu pysähtyy (hyönteiset, linnut, nisäkkäät). Ei kuitenkaan ole terävää rajaa lopullisen ja määrittelemättömän kasvun välillä. Ihminen, nisäkkäät, linnut voivat kasvun pysähtymisen jälkeen vielä hieman kasvaa. Kasvuprosesseja ohjaa genotyyppi, samalla riippuen ympäristöolosuhteista. Ihmisen kasvu, joka johtuu perinnöllisten ja ympäristötekijöiden yhdistelmästä, paljastaa vaihtelua (ikä, sukupuoli, ryhmä, ryhmän sisäinen tai yksilö ja aikakausi). Sen genotyyppi voi myös vaikuttaa epäsuorasti organismin kasvuun ja kehitykseen biologisesti aktiivisten aineiden - hormonien - synteesin kautta. Nämä ovat hermosolujen, endokriinisten rauhasten hormonien, tuottamia hermosareja. Hormonit voivat vaikuttaa sekä aineenvaihduntaprosesseihin (biosynteesiin) että muiden geenien ilmentymiseen, mikä puolestaan vaikuttaa kasvuun. Kaikkien endokriinisten rauhasten välillä on palauteohjattu suhde. Joten aivolisäkkeen hormonit vaikuttavat sukurauhasten, kilpirauhasen ja lisämunuaisten endokriiniseen toimintaan. Aivolisäke tuottaa kasvuhormonia, jonka puute johtaa kääpiöisyyteen - kääpiöisyyteen ja ylimäärä - gigantismiin.
Alkioiden 4. vaihe - lopullisen (lopullisen) organogeneesin vaihe , jolla tapahtuu pysyvien elinten muodostuminen. Tässä alkion synnyn viimeisessä vaiheessa tapahtuvat erittäin monimutkaiset prosessit ovat yksityisen embryologian tutkimuksen kohteena. Tässä osiossa rajoitamme pohtimaan alkion ensisijaisten elinten "kohtaloa".
Ektodermista kehittyvät: ihon orvaskesi ja sen johdannaiset - höyhenet, hiukset, kynnet, iho ja maitorauhaset, hermosto. Hermoputken etuosa (laajennettu) muuttuu aivoiksi, loput (etu- ja keskiosat) selkäytimeksi. Endodermi muodostaa ruuansulatus- ja hengityselinten sisäisen vuorauksen, joka erittää ruuansulatusrauhasten soluja. Somiitit käyvät läpi seuraavat muutokset: dermatomi muodostaa dermiksen (ihon syvän kerroksen); sklerotomi osallistuu luuston (rusto, sitten luun) muodostumiseen; myotomi synnyttää luustolihaksia. Virtsaelimet kehittyvät nefrotomista.
Segmentoimaton mesoderma (splanchnotoma) synnyttää keuhkopussin, vatsakalvon, sydänpussin ja osallistuu sydän- ja verisuonijärjestelmän sekä imukudosjärjestelmän kehitykseen.
Murrosikä - ihmiskehon lisääntymistoimintojen muodostumisprosessi, joka ilmenee toissijaisten seksuaalisten ominaisuuksien asteittaisena kehittymisenä ja päättyy murrosiän alkamiseen. Ihmisillä murrosikää kutsutaan siirtymäkaudeksi eli murrosikää, sen kesto on keskimäärin noin 5 vuotta. Murrosiän ikähaarukka riippuu yksilöllisistä vaihteluista (tytöt 8 - 10 - 16 - 17 vuotta, pojat 10 - 12 - 19 - 20 vuotta). Toissijaisten seksuaalisten ominaisuuksien ilmaantumista 8–10-vuotiailla tytöillä ja 10–12-vuotiailla pojilla kutsutaan varhaiseksi murrosiässä (se liittyy yleensä perustuslailliseen tekijät).
Tärkeä murrosiän merkki - sukurauhasten säännöllisen toiminnan muodostuminen, joka ilmenee tytöillä kuukautisten kautta ja pojilla siemensyöksynä. Sukurauhasten sisäinen eritystoiminta ilmenee molemmilla sukupuolilla myös luuston yksittäisten segmenttien kasvunopeuden faasimuutoksina, minkä seurauksenaruumiin lopulliset (rakenteet) mittasuhteet määritetään ja toissijaisia seksuaalisia ominaisuuksia muodostuu. Toissijaisia seksuaalisia ominaisuuksia ovat pääasiassa muutokset ihossa (erityisesti kivespussissa) ja sen johdannaisissa (kypsymisaikana leijonanharjan kasvu, ns. sukupuolielinten ihon kehittyminen apinoilla ja peuran sarvet ). Ensimmäiset merkit poikien murrosiän kehityksestä sekä kivesten koon kasvu ja kokonaiskasvun kiihtyminen ovat karvojen kasvun voimistuminen ja muutokset kivespussissa. Keskimääräinen ikäjakso yksittäisten merkkien ilmaantumiselle 50 %:lla tutkituista oli: äänimutaatio - 12 vuotta 3,5 kuukautta, häpykarvojen kasvu - 12 vuotta 9,5 kuukautta, kurkunpään kilpirauhasen ruston kasvu - 13 vuotta 3,5 kuukautta, kainalokarvojen kasvu - 13 vuotta 9,5 kuukautta ja kasvojen karvat - 14 vuotta 2 kuukautta Tutkiessaan toissijaisten seksuaalisten ominaisuuksien kestoa ja muodostumisnopeutta V. G. Sidamon Eristavi havaitsi, että yksittäisten murrosiän merkkien kehitysnopeudella on "huippunsa".
Ihmisen lisääntymistoiminto - oman lajinsa lisääntyminen. Ihmisen kyvyn lajina välittää puolet tulevan sukupolven geneettisestä tiedosta isältä äidille takaavat mieskehon lisääntymistoiminnan fysiologiset ominaisuudet. Naiskehon lisääntymistoiminto varmistaa hedelmöitysprosessin, sikiön kohdunsisäisen kehityksen, lapsen syntymän ja sen ruokinnan rintamaidolla. Erottuva ominaisuus henkilön lisääntymistoiminto kehon muista fysiologisista toiminnoista on se, että sen normaali toiminta johtaa uros- ja naarasorganismien sukusolujen fuusioimiseen seksuaalisen lisääntymisen prosessissa. Munasoluja ja siittiöitä kutsutaan naisten ja miesten lisääntymissoluiksi tai sukusoluiksi. Miesten ja naisten sukusolut sisältävät kypsässä muodossaan haploidisen määrän kromosomeja, eli puolet normaalista määrästä. Sukusolujen kromosomien haploidiluku muodostuu spermatogeneesin ja oogeneesin aikana (kuva 16.1). Miehen kehossa spermatogeenisten solujen meioottista jakautumista tapahtuu jatkuvasti koko elämän ajan murrosiän (murrosiän) alkamisen jälkeen. Sitä vastoin munasolussa haploidinen määrä kromosomeja muodostuu välittömästi ennen munan ovulaation alkamista follikkelista. Munasolun ja siittiöiden kyvystä liittyä toisiinsa hedelmöityksen aikana, tsygootin muodostuminen tapahtuu naisen lisääntymiskanavassa. Tätä prosessia kutsutaan lannoitukseksi. Tsygootti sisältää diploidisen määrän kromosomeja, kuten missä tahansa ihmisen ja eläimen kehon somaattisessa solussa. Kaksi kromosomia tsygootin diploidiluvusta, nimittäin sukupuoli-X- ja Y-kromosomit, määräävät uuden sukupolven tulevan yksilön mies- tai naarassukupuolen. Naisen sukusolu sisältää vain X-kromosomit, kun taas miehen sukusolu sisältää X- ja Y-kromosomit. Kromosomit sisältävät geenejä, jotka välittävät geneettisiä ominaisuuksia sukupolvelta toiselle.
Ikääntyminen - tämä on peruuttamaton prosessi, jossa kehon perustoiminnot (regeneratiiviset, lisääntymiskyky jne.) tukahdutetaan asteittain, minkä seurauksena elimistö menettää kykynsä ylläpitää homeostaasia, vastustaa stressiä, sairauksia ja vammoja, mikä tekee kuolemasta väistämättömän .
Kehitysbiologian peruskäsitteet (preformismin ja epigeneesin hypoteesit). Nykyaikaiset käsitteet alkionkehityksen mekanismeista.
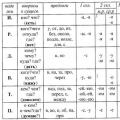 Puheen eri osien päätteiden oikeinkirjoitus
Puheen eri osien päätteiden oikeinkirjoitus Korkeampi ammatillinen koulutus
Korkeampi ammatillinen koulutus Rebus venäjäksi
Rebus venäjäksi