Mendelin toista lakia kutsutaan Mendelin laiksi. Mendelin lakien täyttymisen ehdot. Ominaisuuksien itsenäisen periytymisen laki
Mendelin ensimmäinen laki. Ensimmäisen sukupolven hybridien yhtenäisyyden laki
Kun risteytetään homotsygoottisia yksilöitä, jotka eroavat yhdellä vaihtoehtoisten (toisensa poissulkevien) hahmojen parilla, kaikki jälkeläiset ensimmäinen sukupolvi yhtäläinen sekä fenotyypin että genotyypin suhteen.
Hernekasvien keltaiset (dominoiva ominaisuus) ja vihreät (resessiivinen ominaisuus) siemenet risteytettiin. Sukusolujen muodostumiseen liittyy meioosi. Jokainen kasvi tuottaa yhden tyyppisiä sukusoluja. Jokaisesta homologisesta kromosomiparista yksi kromosomi, jossa on yksi alleelisista geeneistä (A tai a), menee sukusoluihin. Hedelmöityksen jälkeen homologisten kromosomien pariutuminen palautuu ja hybridejä muodostuu. Kaikilla kasveilla on vain keltaisia siemeniä (fenotyyppi), jotka ovat heterotsygoottisia Aa-genotyypille. Tämä tapahtuu, kun täydellinen ylivalta.
Hybridissä Aa on yksi geeni A yhdeltä vanhemmalta ja toinen geeni - a - toiselta vanhemmalta (kuvio 73).
Haploidiset sukusolut (G), toisin kuin diploidiset organismit, on ympyröity.
Risteyksen tuloksena saadaan ensimmäisen sukupolven hybridejä, jotka on merkitty F1.
Risteilyjen kirjaamiseen käytetään erityistä taulukkoa, jota englantilainen geneetikko Punnett ehdotti ja jota kutsutaan Punnett-ruudukoksi.
Isän sukusolut kirjoitetaan vaakasuunnassa ja äidin sukusolut pystysuunnassa. Genotyypitys tallennetaan risteyksissä.

Riisi. 73.Periytys monohybridiristeyksissä.
I - kahden hernelajikkeen risteyttäminen keltaisilla ja vihreillä siemenillä (P); II
Mendelin I ja II lain sytologiset perusteet.
F 1 - heterotsygootit (Aa), F 2 - erottelu genotyypin 1 mukaan AA: 2 Aa: 1 aa.
py jälkeläisiä. Taulukossa solujen määrä riippuu risteyttävien yksilöiden tuottamien sukusolujen määrästä.
Mendelin II laki. Ensimmäisen sukupolven hybridien halkeamislaki
Kun ensimmäisen sukupolven hybridit risteytetään keskenään, toisessa sukupolvessa esiintyy yksilöitä, joilla on sekä hallitsevia että resessiivisiä piirteitä, ja jakautuminen tapahtuu fenotyypin mukaan suhteessa 3:1 (kolme hallitsevaa fenotyyppiä ja yksi resessiivinen) ja 1:2:1. genotyypin mukaan (katso kuva 73). Tällainen jakaminen on mahdollista, kun täydellinen ylivalta.

Hypoteesi sukusolujen "puhtaudesta".
Halkeamislaki voidaan selittää hypoteesilla sukusolujen "puhtaudesta".
Mendel kutsui ilmiötä, jossa vaihtoehtoisten merkkien alleelit eivät sekoitu heterotsygoottisen organismin sukusoluissa (hybridi) hypoteesi sukusolujen "puhtaudesta". Kaksi alleelista geeniä (Aa) on vastuussa kustakin ominaisuudesta. Kun hybridit muodostuvat, alleeliset geenit eivät sekoitu, vaan pysyvät muuttumattomina.
Meioosin seurauksena Aa-hybridit muodostavat kahdenlaisia sukusoluja. Jokainen sukusolu sisältää yhden homologisista kromosomeista, joissa on alleelinen geeni A tai alleelinen geeni a. Sukusolut ovat puhtaita toisesta alleelisesta geenistä. Hedelmöityksen aikana kromosomien homologia ja geenien alleelisuus palautuvat ja ilmaantuu resessiivinen ominaisuus (herneiden vihreä väri), jonka geeni ei osoittanut vaikutustaan hybridiorganismissa. Ominaisuudet kehittyvät geenien vuorovaikutuksen kautta.

Epätäydellinen dominanssi
klo epätäydellinen ylivalta heterotsygoottisilla yksilöillä on oma fenotyyppinsä, ja ominaisuus on keskitasoa.
Kun yökauneuskasveja risteytetään punaisten ja valkoisten kukkien kanssa, ensimmäisessä sukupolvessa ilmestyy vaaleanpunaisia yksilöitä. Ensimmäisen sukupolven hybridien (vaaleanpunaiset kukat) risteyttämisessä jälkeläisten pilkkoutuminen genotyypin ja fenotyypin mukaan osuu yhteen (kuva 74).


Riisi. 74.Perintö, jossa on epätäydellinen dominanssi yökauneuskasvissa.
Geenillä, joka aiheuttaa sirppisoluanemiaa ihmisillä, on epätäydellisen dominanssin ominaisuus.
Analyysi risti
Resessiivinen ominaisuus (vihreät herneet) esiintyy vain homotsygoottisessa tilassa. Homotsygoottiset (keltaherneet) ja heterotsygoottiset (keltaherneet) yksilöt, joilla on hallitsevia ominaisuuksia, eivät eroa toisistaan fenotyypin suhteen, mutta niillä on erilaiset genotyypit. Niiden genotyypit voidaan määrittää risteyttämällä yksilöiden kanssa, joilla on tunnettu genotyyppi. Tällainen yksilö voi olla vihreitä herneitä, joilla on homotsygoottinen resessiivinen ominaisuus. Tätä ristiä kutsutaan analysoitavaksi ristiksi. Jos risteyttämisen seurauksena kaikki jälkeläiset ovat yhtenäisiä, tutkittava yksilö on homotsygoottinen.
Jos jakautuminen tapahtuu, yksilö on heterotsygoottinen. Heterotsygoottisen yksilön jälkeläiset pilkkoutuvat suhteessa 1:1.

Mendelin III laki. Ominaisuuksien itsenäisen yhdistelmän laki (Kuva 75). Organismit eroavat toisistaan monella tapaa.
Kahden ominaisuuden suhteen eroavien yksilöiden risteytymistä kutsutaan dihybridiksi ja monessa suhteessa polyhybridiksi.
Kun risteytetään homotsygoottisia yksilöitä, jotka eroavat kahdella vaihtoehtoisella hahmoparilla, tapahtuu toisessa sukupolvessa itsenäinen ominaisuuksien yhdistelmä.
Dihybridiristeytyksen seurauksena koko ensimmäinen sukupolvi on yhtenäinen. Toisessa sukupolvessa fenotyyppinen pilkkoutuminen tapahtuu suhteessa 9:3:3:1.
Jos esimerkiksi ristät herneen, jossa on keltaiset siemenet ja sileä pinta (dominoiva ominaisuus) herneen kanssa, jossa on vihreitä siemeniä ja ryppyinen pinta (resessiivinen ominaisuus), koko ensimmäinen sukupolvi on yhtenäinen (keltaiset ja sileät siemenet).


Kun hybridit risteytettiin keskenään toisessa sukupolvessa, ilmaantui yksilöitä, joilla oli ominaisuuksia, joita ei ollut alkuperäisissä muodoissa (keltaiset ryppyiset ja vihreät sileät siemenet). Nämä ominaisuudet ovat periytyviä riippumatta toisiltaan.

Diheterotsygoottinen yksilö tuotti 4 tyyppiä sukusoluja
Hybridien risteyttämisen jälkeen toisen sukupolven syntyneiden yksilöiden laskemisen helpottamiseksi käytetään Punnett-ruudukkoa.


Riisi. 75.Ominaisuuksien itsenäinen jakautuminen dihybridiristeyksissä. A, B, a, b - hallitsevat ja resessiiviset alleelit, jotka ohjaavat kahden ominaisuuden kehitystä. G - vanhempien sukusolut; F 1 - ensimmäisen sukupolven hybridit; F 2 - toisen sukupolven hybridit.
Meioosin seurauksena jokaiseen sukusoluun siirtyy yksi alleelisista geeneistä homologisesta kromosomiparista.
Muodostuu 4 tyyppiä sukusoluja. Katkaisu risteytyksen jälkeen suhteessa 9:3:3:1 (9 yksilöä, joilla on kaksi hallitsevaa piirrettä, 1 yksilö, jolla on kaksi resessiivistä ominaisuutta, 3 yksilöä, joilla on yksi dominantti ja toiset resessiiviset piirteet, 3 yksilöä, joilla on dominantti ja resessiivinen piirre).
Dominantteja ja resessiivisiä piirteitä omaavien yksilöiden ilmaantuminen on mahdollista, koska herneiden väristä ja muodosta vastaavat geenit sijaitsevat erilaisissa ei-homologisissa kromosomeissa.
Jokainen alleelisten geenien pari jakautuu toisesta parista riippumatta, ja siksi geenejä voidaan yhdistää itsenäisesti.
Heterotsygoottinen yksilö "n" ominaisuusparille muodostaa 2 n tyyppistä sukusolua.
Kysymyksiä itsehillintää varten
1. Miten Mendelin ensimmäinen laki on muotoiltu?
2. Mitä siemeniä Mendel risteili herneiden kanssa?
3. Mitä siemeniä sisältävät kasvit ovat syntyneet risteyttämisestä?
4. Miten Mendelin II laki on muotoiltu?
5. Mitä ominaisuuksia omaavia kasveja saatiin ensimmäisen sukupolven hybridien risteyttämisen tuloksena?
6. Millä numeerisella suhteella jakautuminen tapahtuu?
7. Miten jakamisen laki voidaan selittää?
8. Kuinka selittää hypoteesi sukusolujen "puhtaudesta"?
9. Miten selittää piirteiden epätäydellinen dominanssi? 10. Millaista katkaisua fenotyypin ja genotyypin mukaan tapahtuu?
ensimmäisen sukupolven hybridien risteyttämisen jälkeen?
11. Milloin analyyttinen risti suoritetaan?
12. Miten analyyttinen risti suoritetaan?
13. Millaista ristiä kutsutaan dihybridiksi?
14. Missä kromosomeissa herneiden väristä ja muodosta vastaavat geenit sijaitsevat?
15. Miten Mendelin III laki on muotoiltu?
16. Mikä fenotyyppinen pilkkoutuminen tapahtuu ensimmäisessä sukupolvessa?
17. Millaista fenotyyppistä pilkkoutumista tapahtuu toisessa sukupolvessa?
18. Mitä käytetään hybridien risteytyksen tuloksena syntyneiden yksilöiden laskemisen helpottamiseksi?
19.Miten voimme selittää yksilöiden ulkonäön, joilla on ominaisuuksia, joita ei ennen ollut?
Aiheen "Mendelin lait" avainsanat
alleelisuusanemia
vuorovaikutusta
sukusolut
geeni
genotyyppi
heterotsygootti
hybridi
hypoteesi sukusolujen "puhtaudesta".
homotsygootti
homologiaa
herneet
herne
toiminta
dihybridi
valta-asema
yhtenäisyys
laki
meioosi
koulutus väritys
lannoitus
yksilöllinen
pariliitoksen muodostaminen
pinta
Kreivi
sukupolvi
polyhybridi
jälkeläisiä
ulkomuoto
merkki
tehdas
jakaa
Punett-ruudukko
vanhemmat
omaisuutta
siemenet
ylitys
fuusio
suhde
lajike
sopivuus
fenotyyppi
muodossa
merkki
väri
kukat
Monikertainen alleelismi
Alleeliset geenit eivät voi sisältää kahta, vaan suuremman määrän geenejä. Nämä ovat useita alleeleja. Ne syntyvät mutaation seurauksena (DNA-molekyylin nukleotidin korvaaminen tai häviäminen). Esimerkki useista alleeleista voivat olla ihmisen veriryhmistä vastaavat geenit: I A, I B, I 0. Geenit I A ja IB ovat hallitsevia I 0-geenille. Genotyypissä on aina vain kaksi geeniä alleelisarjasta. Geenit I 0 I 0 määrittävät veriryhmän I, geenit I A I A, I A I O - ryhmä II, I B I B, I B I 0 - ryhmä III, I A I B - ryhmä IV.
Geenien vuorovaikutus
Geenin ja ominaisuuden välillä on monimutkainen suhde. Yksi geeni voi olla vastuussa yhden ominaisuuden kehittymisestä.
Geenit ovat vastuussa proteiinien synteesistä, jotka katalysoivat tiettyjä biokemiallisia reaktioita, jotka johtavat tiettyihin ominaisuuksiin.
Yksi geeni voi olla vastuussa useiden ominaisuuksien kehittymisestä pleiotrooppinen vaikutus. Geenin pleiotrooppisen vaikutuksen vakavuus riippuu biokemiallisesta reaktiosta, jonka katalysoi entsyymi, joka syntetisoituu tämän geenin ohjauksessa.
Useat geenit voivat olla vastuussa yhden piirteen kehittymisestä - tämä on polymeeri geenin toiminta.
Oireiden ilmeneminen on seurausta erilaisten biokemiallisten reaktioiden vuorovaikutuksesta. Nämä vuorovaikutukset voidaan yhdistää alleelisiin ja ei-alleelisiin geeneihin.
Alleelisten geenien vuorovaikutus.
Samassa alleeliparissa sijaitsevien geenien vuorovaikutus tapahtuu seuraavasti:
. täydellinen määräävä asema;
. epätäydellinen määräävä asema;
. yhteisvalta-asema;
. ylivaltaa.
klo saattaa loppuun Dominanssissa yhden (dominoivan) geenin toiminta tukahduttaa täysin toisen (resessiivisen) toiminnan. Risteytyksen yhteydessä vallitseva ominaisuus ilmenee ensimmäisessä sukupolvessa (esimerkiksi herneiden keltainen väri).
klo epätäydellinen Dominanssi syntyy, kun hallitsevan alleelin vaikutus heikkenee resessiivisen alleelin läsnä ollessa. Risteyksen tuloksena saaduilla heterotsygoottisilla yksilöillä on oma genotyyppinsä. Esimerkiksi risteyttäessä yökauneuskasveja punaisten ja valkoisten kukkien kanssa, näkyviin tulee vaaleanpunaisia kukkia.
klo yhteisvalta-asema Molempien geenien vaikutus ilmenee, kun ne ovat läsnä samanaikaisesti. Seurauksena on uusi oire.
Esimerkiksi ihmisen veriryhmä IV (I A I B) muodostuu geenien I A ja I B vuorovaikutuksesta. Erikseen I A-geeni määrittää II-veriryhmän ja IB-geeni III-veriryhmän.
klo ylivaltaa Heterotsygoottisessa tilassa hallitsevalla alleelilla on vahvempi ilmentymä piirteestä kuin homotsygoottisessa tilassa.
Ei-alleelisten geenien vuorovaikutus
Organismin yhteen ominaisuuteen voivat usein vaikuttaa useat ei-alleelisten geenien parit.
Ei-alleelisten geenien vuorovaikutus tapahtuu seuraavasti:
. täydentävyys;
. epistaasi;
. polymeerit.
Täydentävä vaikutus ilmenee kahden hallitsevan ei-alleelisen geenin samanaikaisella läsnäololla organismien genotyypissä. Kumpikin hallitsevista geeneistä voi ilmaantua itsenäisesti, jos toinen on resessiivisessä tilassa, mutta niiden yhteinen esiintyminen dominoivassa tilassa tsygootissa määrittää ominaisuuden uuden tilan.
Esimerkki. Kaksi valkokukkaisten herneiden lajiketta risteytettiin. Kaikilla ensimmäisen sukupolven hybrideillä oli punaiset kukat. Kukkien väri riippuu kahdesta vuorovaikutuksessa olevasta geenistä A ja B.

Geenien A ja B perusteella syntetisoidut proteiinit (entsyymit) katalysoivat biokemiallisia reaktioita, jotka johtavat ominaisuuden (kukkien punaisen värin) ilmenemiseen.
Epistasis- vuorovaikutus, jossa yksi hallitsevista tai resessiivisistä ei-alleelisista geeneistä suppressoi toisen ei-alleelisen geenin toimintaa. Geeniä, joka vaimentaa toisen toimintaa, kutsutaan epistaattiseksi geeniksi tai suppressoriksi. Tukahdutettua geeniä kutsutaan hypostaattiseksi. Epistasis voi olla hallitseva tai resessiivinen.
Hallitseva epistaasi. Esimerkki hallitsevasta epistaasista olisi höyhenen värin periytyminen kanoissa. Hallitseva geeni C on vastuussa höyhenen väristä. Hallitseva ei-alleelinen geeni I estää höyhenen värin kehittymisen. Tämän seurauksena kanoilla, joiden genotyypissä on C-geeni, on I-geenin läsnä ollessa valkoinen höyhenpeite: IICC; IICC; IiCc; Iicc. Kanat, joilla on genotyyppi iicc, ovat myös valkoisia, koska nämä geenit ovat resessiivisessä tilassa. Kanojen höyhenet, joilla on genotyyppi iiCC, iiCc, ovat värillisiä. Höyhenen valkoinen väri johtuu i-geenin resessiivisestä alleelista tai väriä suppressorigeenin I läsnäolosta. Geenien vuorovaikutus perustuu entsyymiproteiinien välisiin biokemiallisiin yhteyksiin, joita epistaattiset geenit koodaavat.
Resessiivinen epistaasi. Resessiivinen epistaasi selittää Bombay-ilmiön - ABO-veriryhmäjärjestelmän antigeenien epätavallisen periytymisen. Veriryhmiä tunnetaan 4.
Veriryhmän I (I 0 I 0) naisen perheeseen II veriryhmän (I A I A) mies synnytti IV-veriryhmän (I A I B) lapsen, mikä on mahdotonta. Kävi ilmi, että nainen peri I B-geenin äidiltään ja I 0 -geenin isältään. Vain I 0 -geeni osoitti siis vaikutuksen
naisen uskottiin olevan veriryhmä I. Geeni I B tukahdutti resessiivisen geenin x, joka oli homotsygoottisessa tilassa - xx.

Tämän naisen lapsessa tukahdutettu I B-geeni osoitti vaikutuksensa. Lapsella oli IV veriryhmä I A I B.
PolymeeriGeenien vaikutus johtuu siitä, että useat ei-alleeliset geenit voivat olla vastuussa samasta ominaisuudesta, mikä tehostaa sen ilmenemistä. Ominaisuudet, jotka riippuvat polymeerigeeneistä, luokitellaan kvantitatiivisiksi. Kvantitatiivisten ominaisuuksien kehittymisestä vastuussa olevilla geeneillä on kumulatiivinen vaikutus. Esimerkiksi polymeeriset ei-alleeliset geenit S1 ja S2 ovat vastuussa ihon pigmentaatiosta ihmisillä. Näiden geenien hallitsevien alleelien läsnä ollessa syntetisoituu paljon pigmenttiä, resessiivisten läsnä ollessa - vähän. Ihon värin voimakkuus riippuu pigmentin määrästä, jonka määrää hallitsevien geenien määrä.

Mulattien S 1 s 1 S 2 s 2 avioliitosta lapsilla on ihon pigmentti vaaleasta tummaan, mutta todennäköisyys saada lapsi, jolla on valkoinen ja musta iho, on 1/16.
Monet ominaisuudet periytyvät polymeeriperiaatteen mukaisesti.
Kysymyksiä itsehillintää varten
1. Mitä ovat useat alleelit?
2. Mitkä geenit ovat vastuussa ihmisen verityypeistä?
3. Mitä veriryhmiä ihmisellä on?
4. Mitä yhteyksiä geenin ja ominaisuuden välillä on?
5. Miten alleeliset geenit ovat vuorovaikutuksessa?
6. Miten ei-alleeliset geenit ovat vuorovaikutuksessa?
7. Miten geenin täydentävä vaikutus voidaan selittää?
8. Miten epistaasi voidaan selittää?
9. Miten geenin polymeerinen vaikutus voidaan selittää?
Aiheen "Useita alleeleja ja geenivuorovaikutus" avainsanat
alleelismi alleeli antigeenit avioliitto
vuorovaikutusta
genotyyppi
hybridi
herneet
herneet
veriryhmä
toiminta
lapset
valta-asema
nainen
korvaus
kodominanssi
yhteisvalta-asema
nahka
kanat
äiti
molekyyli
mulatti
mutaatio
Saatavuus
perinnöstä
nukleotidit
väritys
höyhenpeite
perusta
asenne
pigmentti
pigmentaatio
pleiotropia
vaimennin
sukupolvi
polymerismi
merkki
esimerkki
läsnäolo
ilmentymä
kehitystä
reaktiot
lapsi
tulos
ylivalta-yhteys
proteiinisynteesijärjestelmä
ylitys
osavaltio
tutkinnon
menetys
ilmiö
entsyymejä
väri
kukat
Ihmisen
Genetiikka- tiede perinnöllisyyden ja vaihtelevuuden laeista. Genetiikan ”syntymisajankohdaksi” voidaan katsoa vuotta 1900, jolloin G. De Vries Hollannissa, K. Correns Saksassa ja E. Cermak Itävallassa itsenäisesti ”löysivät” G. Mendelin vuonna 1999 määrittelemät ominaisuuksien periytymislait. 1865.
Perinnöllisyys- eliöiden kyky välittää ominaisuuksiaan sukupolvelta toiselle.
Vaihtuvuus- eliöiden ominaisuus hankkia uusia ominaisuuksia vanhempiinsa verrattuna. Laajassa merkityksessä vaihtelu viittaa eroihin saman lajin yksilöiden välillä.
Merkki- mikä tahansa rakenteellinen piirre, mikä tahansa kehon ominaisuus. Ominaisuuden kehittyminen riippuu sekä muiden geenien läsnäolosta että ympäristöolosuhteista. Siksi jokaisella yksilöllä on joukko ominaisuuksia, jotka ovat ominaisia vain hänelle.
Fenotyyppi- kehon kaikkien ulkoisten ja sisäisten merkkien kokonaisuus.
Gene- toiminnallisesti jakamaton geneettisen materiaalin yksikkö, DNA-molekyylin osa, joka koodaa polypeptidin, siirto- tai ribosomaalisen RNA-molekyylin primäärirakennetta. Laajassa mielessä geeni on DNA:n osa, joka määrittää mahdollisuuden kehittää erillinen perusominaisuus.
Genotyyppi- organismin geenien joukko.
Locus- geenin sijainti kromosomissa.
Alleeliset geenit- geenit, jotka sijaitsevat homologisten kromosomien identtisissä lokuksissa.
Homotsygootti- organismi, jolla on yhden molekyylimuodon alleelisia geenejä.
Heterotsygootti- organismi, jolla on eri molekyylimuotoisia alleelisia geenejä; tässä tapauksessa yksi geeneistä on hallitseva, toinen on resessiivinen.
Resessiivinen geeni- alleeli, joka määrittää piirteen kehittymisen vain homotsygoottisessa tilassa; tällaista ominaisuutta kutsutaan resessiiviseksi.
Dominoiva geeni- alleeli, joka määrittää ominaisuuden kehittymisen ei vain homotsygoottisessa, vaan myös heterotsygoottisessa tilassa; tällaista ominaisuutta kutsutaan hallitsevaksi.
Geneettiset menetelmät
Pääasiallinen on hybridologinen menetelmä- risteysjärjestelmä, jonka avulla voidaan jäljittää piirteiden periytymismalleja useiden sukupolvien aikana. Ensimmäiseksi kehitti ja käytti G. Mendel. Menetelmän tunnusmerkit: 1) kohdennettu valinta vanhemmista, jotka eroavat yhden, kahden, kolmen jne. vastakkaisten (vaihtoehtoisten) stabiilien ominaisuuksien parin suhteen; 2) hybridien ominaisuuksien periytymisen tiukka määrällinen laskenta; 3) kunkin vanhemman jälkeläisten yksilöllinen arviointi sukupolvien sarjassa.
Risteystä, jossa analysoidaan yhden vaihtoehtoisen merkkiparin periytymistä, kutsutaan monohybridi, kaksi paria - dihybridi, useita pareja - polyhybridi. Vaihtoehtoisilla ominaisuuksilla tarkoitetaan piirteen erilaisia merkityksiä, esimerkiksi piirre on herneiden väri, vaihtoehtoiset ominaisuudet ovat keltainen väri, herneiden vihreä väri.
Hybridologisen menetelmän lisäksi genetiikassa käytetään seuraavia: sukututkimus— sukutaulujen kokoaminen ja analysointi; sytogeneettinen— kromosomien tutkimus; kaksos— kaksosten tutkimus; väestötilastollinen menetelmä - populaatioiden geneettisen rakenteen tutkiminen.
Geneettinen symboliikka
G. Mendelin ehdotus, käytetään risteytysten tulosten kirjaamiseen: P - vanhemmat; F - jälkeläinen, numero kirjaimen alla tai välittömästi sen jälkeen osoittaa sukupolven sarjanumeron (F 1 - ensimmäisen sukupolven hybridit - vanhempien suorat jälkeläiset, F 2 - toisen sukupolven hybridit - syntyvät F 1 -hybridien risteyttämisen seurauksena jokaisen kanssa muu); × — risteyskuvake; G - mies; E - nainen; A on hallitseva geeni, a on resessiivinen geeni; AA on homotsygootti dominantille, aa on homotsygootti resessiiviselle, Aa on heterotsygootti.
Ensimmäisen sukupolven hybridien yhtenäisyyslaki tai Mendelin ensimmäinen laki
Mendelin työn onnistumista edesauttoi onnistunut risteytyskohteen valinta - eri hernelajikkeet. Herneiden ominaisuudet: 1) se on suhteellisen helppo kasvattaa ja sillä on lyhyt kehitysjakso; 2) hänellä on useita jälkeläisiä; 3) sillä on suuri määrä selvästi näkyviä vaihtoehtoisia ominaisuuksia (terän väri - valkoinen tai punainen; sirkkalehtien väri - vihreä tai keltainen; siemenen muoto - ryppyinen tai sileä; palkojen väri - keltainen tai vihreä; palkon muoto - pyöreä tai ahtautunut; kukka-asetelma tai hedelmät - koko varren pituudella tai sen yläosassa - pitkä tai lyhyt); 4) on itsepölyttäjä, minkä seurauksena sillä on suuri määrä puhtaita linjoja, jotka säilyttävät ominaisuutensa vakaasti sukupolvelta toiselle.
Mendel teki kokeita eri hernelajikkeiden risteyttämisestä kahdeksan vuoden ajan vuodesta 1854 alkaen. Helmikuun 8. päivänä 1865 G. Mendel puhui Brunn Society of Naturalists -järjestön kokouksessa raportilla "Experiments on Experiments on kasvi hybrids", jossa hänen työnsä tuloksista tehtiin yhteenveto.
Mendelin kokeet harkittiin huolellisesti. Jos hänen edeltäjänsä yrittivät tutkia useiden piirteiden periytymismalleja kerralla, Mendel aloitti tutkimuksensa tutkimalla vain yhden vaihtoehtoisen piirteen periytymistä.
Mendel otti hernelajikkeita, joissa oli keltaisia ja vihreitä siemeniä, ja ristipölytti ne keinotekoisesti: hän poisti heteet yhdestä lajikkeesta ja pölytti ne toisen lajikkeen siitepölyllä. Ensimmäisen sukupolven hybrideissä oli keltaiset siemenet. Samanlainen kuva havaittiin risteyksissä, joissa tutkittiin muiden ominaisuuksien periytymistä: risteytettäessä kasveja, joilla oli sileä ja ryppyinen siemenmuoto, kaikki syntyneiden hybridien siemenet olivat sileitä, kun risteytettiin punakukkaisia kasveja valkokukkaisten kasvien kanssa tuloksena saadut olivat punakukkaisia. Mendel tuli siihen tulokseen, että ensimmäisen sukupolven hybrideissä kustakin vaihtoehtoisten hahmojen parista vain yksi ilmestyy ja toinen näyttää katoavan. Mendel kutsui ensimmäisen sukupolven hybrideissä ilmenevää ominaisuutta hallitsevaksi ja tukahdutettua ominaisuutta resessiiviseksi.
klo homotsygoottisten yksilöiden monohybridiristeytys hybrideillä on erilaisia vaihtoehtoisten ominaisuuksien arvoja, ja ne ovat genotyypiltään ja fenotyypiltään yhdenmukaisia.
Mendelin yhtenäisyyslain geneettinen kaavio
(A on herneiden keltainen väri ja herneiden vihreä väri)
Erottelulaki tai Mendelin toinen laki
G. Mendel antoi ensimmäisen sukupolven hybrideille mahdollisuuden itsepölytykseen. Tällä tavalla saadut toisen sukupolven hybridit osoittivat paitsi hallitsevan myös resessiivisen ominaisuuden. Kokeiden tulokset on esitetty taulukossa.
| Merkkejä | Hallitseva | Resessiivinen | Kaikki yhteensä | ||
|---|---|---|---|---|---|
| Määrä | % | Määrä | % | ||
| Siemenen muoto | 5474 | 74,74 | 1850 | 25,26 | 7324 |
| Sirkkalehtien väri | 6022 | 75,06 | 2001 | 24,94 | 8023 |
| Siemenkuoren väri | 705 | 75,90 | 224 | 24,10 | 929 |
| Bobin muoto | 882 | 74,68 | 299 | 25,32 | 1181 |
| Bob väritys | 428 | 73,79 | 152 | 26,21 | 580 |
| Kukka-asetelma | 651 | 75,87 | 207 | 24,13 | 858 |
| Varren korkeus | 787 | 73,96 | 277 | 26,04 | 1064 |
| Kaikki yhteensä: | 14949 | 74,90 | 5010 | 25,10 | 19959 |
Taulukon tietojen analysointi antoi meille mahdollisuuden tehdä seuraavat johtopäätökset:
- Toisessa sukupolvessa hybridit eivät ole yhdenmukaisia: joissakin hybrideissä on yksi (dominoiva), toisissa - toinen (resessiivinen) piirre vaihtoehtoisesta parista;
- hallitsevan ominaisuuden sisältävien hybridien määrä on noin kolme kertaa suurempi kuin resessiivisen ominaisuuden sisältävien hybridien lukumäärä;
- Resessiivinen ominaisuus ei katoa ensimmäisen sukupolven hybrideissä, vaan se vain tukahdutetaan ja ilmenee toisessa hybridisukupolvessa.
Ilmiö, jossa osa toisen sukupolven hybrideistä kantaa hallitsevaa ominaisuutta ja osa - resessiivistä, on ns. jakaminen. Lisäksi hybrideissä havaittu halkeaminen ei ole satunnaista, vaan siihen liittyy tiettyjä määrällisiä malleja. Tämän perusteella Mendel teki toisen johtopäätöksen: ensimmäisen sukupolven hybridejä risteyttäessä jälkeläisten ominaisuudet jaetaan tietyssä numeerisessa suhteessa.
klo heterotsygoottisten yksilöiden monohybridiristeytys hybrideissä tapahtuu katkaisu fenotyypin mukaan suhteessa 3:1, genotyypin mukaan 1:2:1.
Mendelin erottelulain geneettinen kaavio
(A on herneiden keltainen väri ja herneiden vihreä väri):
Sukusolujen puhtauden laki
Vuodesta 1854, kahdeksan vuoden ajan, Mendel suoritti kokeita hernekasvien risteyttämisestä. Hän havaitsi, että eri hernelajikkeiden risteyttämisen seurauksena ensimmäisen sukupolven hybrideillä on sama fenotyyppi, ja toisen sukupolven hybrideissä ominaisuudet jakautuvat tietyissä suhteissa. Tämän ilmiön selittämiseksi Mendel teki useita oletuksia, joita kutsuttiin "sukusolun puhtaushypoteesiksi" tai "sukusolun puhtauslakiksi". Mendel ehdotti seuraavaa:
- jotkin erilliset perinnölliset tekijät ovat vastuussa ominaisuuksien muodostumisesta;
- organismit sisältävät kaksi tekijää, jotka määräävät ominaisuuden kehittymisen;
- sukusolujen muodostumisen aikana vain yksi tekijäparista tulee jokaiseen;
- kun uros- ja naaraspuoliset sukusolut sulautuvat yhteen, nämä perinnölliset tekijät eivät sekoitu (pysyvät puhtaina).
Vuonna 1909 V. Johansen kutsui näitä perinnöllisiä tekijöitä geeneiksi, ja vuonna 1912 T. Morgan osoitti, että ne sijaitsevat kromosomeissa.
Todistaakseen olettamuksiaan G. Mendel käytti ylitystä, jota nykyään kutsutaan analysoimiseksi ( testi risti- tuntemattoman genotyypin organismin risteyttäminen resessiiviselle homotsygoottiselle organismille). Mendel luultavasti päätteli seuraavasti: "Jos oletukseni ovat oikeat, risteyttämisen seurauksena F 1 -lajikkeen kanssa, jolla on resessiivinen ominaisuus (vihreät herneet), hybridien joukossa on puoliksi vihreitä ja puoliksi keltaisia herneitä." Kuten alla olevasta geneettisestä kaaviosta voidaan nähdä, hän itse asiassa jakoi 1:1 ja oli vakuuttunut olettamustensa ja päätelmiensä oikeellisuudesta, mutta hänen aikalaisensa eivät ymmärtäneet häntä. Hänen Brunn Society of Naturalists -yhdistyksen kokouksessa tehty raportti "Kasvien hybridien kokeet" otettiin vastaan täydellisellä hiljaisuudella.
Mendelin ensimmäisen ja toisen lain sytologinen perusta
Mendelin aikaan sukusolujen rakennetta ja kehitystä ei ollut tutkittu, joten hänen hypoteesinsa sukusolujen puhtaudesta on esimerkki loistavasta ennakoinnista, joka sai myöhemmin tieteellisen vahvistuksen.
Mendelin havaitsemat dominanssi- ja hahmojen erotteluilmiöt selittyvät tällä hetkellä kromosomien paristumisella, kromosomien poikkeavilla meioosin aikana ja niiden yhdistymisellä hedelmöittymisen aikana. Merkitään keltaisen värin määräävää geeniä kirjaimella A ja vihreää a:lla. Koska Mendel työskenteli puhtaiden linjojen kanssa, molemmat risteytetyt organismit ovat homotsygoottisia, eli niillä on kaksi identtistä siemenvärigeenin alleelia (AA ja aa, vastaavasti). Meioosin aikana kromosomien määrä puolittuu, ja vain yksi kromosomi parista päätyy jokaiseen sukusoluun. Koska homologisissa kromosomeissa on samat alleelit, yhden organismin kaikki sukusolut sisältävät kromosomin, jossa on geeni A, ja toisen - geenin a.
Hedelmöityksen aikana uros- ja naarassukusolut sulautuvat yhteen ja niiden kromosomit yhdistyvät yhdeksi tsygootiksi. Tuloksena oleva hybridi tulee heterotsygoottiseksi, koska sen soluilla on Aa-genotyyppi; yksi genotyypin variantti antaa yhden fenotyypin variantin - herneiden keltaisen värin.
Hybridiorganismissa, jolla on Aa-genotyyppi meioosin aikana, kromosomit erottuvat eri soluiksi ja muodostuu kahdenlaisia sukusoluja - puolet sukusoluista kantaa geeniä A, toinen puoli kantaa geeniä a. Hedelmöityminen on satunnainen ja yhtä todennäköinen prosessi, eli mikä tahansa siittiö voi hedelmöittää minkä tahansa munasolun. Koska muodostui kahdenlaisia siittiöitä ja kaksi tyyppiä munasoluja, neljä tsygoottityyppiä on mahdollista. Puolet heistä on heterotsygootteja (kannattaa A- ja a-geeniä), 1/4 on homotsygoottisia hallitsevan ominaisuuden suhteen (kantaa A-geeniä) ja 1/4 on homotsygoottisia resessiivisen ominaisuuden suhteen (kaksi a-geeniä). Homotsygootit hallitseville ja heterotsygooteille tuottavat keltaisia herneitä (3/4), homotsygootit resessiivisille - vihreitä (1/4).
Ominaisuuksien itsenäisen yhdistämisen (perinnön) laki tai Mendelin kolmas laki
Organismit eroavat toisistaan monella tapaa. Sen vuoksi saatuaan yhden piirreparin periytymismallit G. Mendel siirtyi tutkimaan kahden (tai useamman) vaihtoehtoisten ominaisuusparien periytymistä. Dihybridiristeyksissä Mendel otti homotsygoottisia hernekasveja, jotka erosivat siementen väriltään (keltainen ja vihreä) ja siemenen muodosta (sileät ja ryppyiset). Siementen keltainen väri (A) ja sileä muoto (B) ovat hallitsevia merkkejä, vihreä väri (a) ja ryppyinen muoto (b) ovat resessiivisiä merkkejä.
Ristittämällä keltaisia ja sileitä siemeniä sisältävän kasvin vihreitä ja ryppyisiä siemeniä sisältävän kasvin kanssa Mendel sai yhtenäisen hybridisukupolven F 1 keltaisilla ja sileillä siemenillä. 15 ensimmäisen sukupolven hybridin itsepölytyksestä saatiin 556 siementä, joista 315 oli keltaisia sileitä, 101 keltaryppyisiä, 108 vihreää sileitä ja 32 vihreää ryppyisiä (halkaisu 9:3:3:1).
Analysoidessaan tuloksena saatuja jälkeläisiä Mendel kiinnitti huomion siihen, että: 1) alkuperäisten lajikkeiden ominaisuuksien yhdistelmien (keltaiset sileät ja vihreät ryppyiset siemenet) rinnalla dihybridiristeytyksen aikana ilmaantuu uusia ominaisuuksien yhdistelmiä (keltaiset ryppyiset ja vihreät sileät siemenet); 2) jakautuminen kunkin yksittäisen ominaisuuden osalta vastaa jakautumista monohybridiristeytyksen aikana. 556 siemenestä 423 oli sileitä ja 133 ryppyisiä (suhde 3:1), 416 siemeniä oli väriltään keltaisia ja 140 vihreitä (suhde 3:1). Mendel tuli siihen tulokseen, että jakautuminen yhdessä ominaisuusparissa ei liity jakautumiseen toisessa parissa. Hybridisiemenille ei ole ominaista vain emokasvien ominaisuuksien yhdistelmät (keltaiset sileät siemenet ja vihreät ryppyiset siemenet), vaan myös uusien ominaisuuksien yhdistelmien ilmaantuminen (keltaiset ryppyiset siemenet ja vihreät sileät siemenet).
Kun dihybridi risteyttää diheterotsygootteja hybrideissä, tapahtuu katkaisu fenotyypin mukaan suhteessa 9:3:3:1, genotyypin mukaan suhteessa 4:2:2:2:2:1:1:1:1. , merkit periytyvät toisistaan riippumatta ja yhdistetään kaikkiin mahdollisiin yhdistelmiin.
| R | ♀AABB keltainen, sileä |
× | ♂aabb vihreä, ryppyinen |
| Sukusolujen tyypit | AB | ab | |
| F 1 | AaBb keltainen, sileä, 100% |
||
| P | ♀ AaBb keltainen, sileä |
× | ♂ AABb keltainen, sileä |
| Sukusolujen tyypit | AB Ab aB ab | AB Ab aB ab | |
Piirteiden itsenäisen yhdistelmän lain geneettinen kaavio:
| Sukusolut: | ♂ | AB | Ab | aB | ab |
| ♀ | |||||
| AB | AABB keltainen sileä |
AABb keltainen sileä |
AaBB keltainen sileä |
AaBb keltainen sileä |
|
| Ab | AABb keltainen sileä |
AAbb keltainen ryppyinen |
AaBb keltainen sileä |
Aabb keltainen ryppyinen |
|
| aB | AaBB keltainen sileä |
AaBb keltainen sileä |
aaBB vihreä sileä |
aaBb vihreä sileä |
|
| ab | AaBb keltainen sileä |
Aabb keltainen ryppyinen |
aaBb vihreä sileä |
aabb vihreä ryppyinen |
|
Risteytystulosten analyysi fenotyypin mukaan: keltainen, sileä - 9/16, keltainen, ryppyinen - 3/16, vihreä, sileä - 3/16, vihreä, ryppyinen - 1/16. Fenotyypin jakautuminen on 9:3:3:1.
Risteytystulosten analyysi genotyypin mukaan: AaBb - 4/16, AABb - 2/16, AaBB - 2/16, Aabb - 2/16, aaBb - 2/16, AABB - 1/16, Aabb - 1/16, aaBB - 1/16, aabb - 1/16. Genotyypin jakautuminen on 4:2:2:2:2:1:1:1:1.
Jos monohybridiristeyksessä emoorganismit eroavat yhdellä merkkiparilla (keltaiset ja vihreät siemenet) ja antavat toisessa sukupolvessa kaksi fenotyyppiä (2 1) suhteessa (3 + 1) 1, niin dihybridissä ne eroavat kahdella. merkkipareja ja anna toisessa sukupolvessa neljä fenotyyppiä (2 2) suhteessa (3 + 1) 2. On helppo laskea, kuinka monta fenotyyppiä ja missä suhteessa muodostuu toisessa sukupolvessa trihybridiristityksen aikana: kahdeksan fenotyyppiä (2 3) suhteessa (3 + 1) 3.
Jos genotyypin jakautuminen F 2:ssa monohybridisukupolvella oli 1:2:1, eli oli kolme eri genotyyppiä (3 1), niin dihybridiristeytyksellä muodostuu 9 eri genotyyppiä - 3 2, trihybridiristeytyksellä Muodostuu 3 3 - 27 erilaista genotyyppiä.
Mendelin kolmas laki pätee vain niissä tapauksissa, joissa analysoitujen ominaisuuksien geenit sijaitsevat eri homologisissa kromosomipareissa.
Mendelin kolmannen lain sytologinen perusta
Olkoon A geeni, joka määrää siementen keltaisen värin kehittymisen, a - vihreä väri, B - siemenen sileä muoto, b - ryppyinen. Ensimmäisen sukupolven hybridit, joiden genotyyppi on AaBb, risteytetään. Sukusolujen muodostumisen aikana jokaisesta alleelisten geenien parista vain yksi pääsee sukusoluun ja kromosomien satunnaisen poikkeaman seurauksena meioosin ensimmäisessä jakautumisessa geeni A voi päätyä samaan sukusoluun geenin B tai geenin kanssa. b, ja geeni a - geenin B tai geenin b kanssa. Siten jokainen organismi tuottaa neljää sukusolutyyppiä samassa määrässä (25%): AB, Ab, aB, ab. Hedelmöityksen aikana jokainen neljästä siittiötyypistä voi hedelmöittää minkä tahansa neljästä munatyypistä. Hedelmöityksen seurauksena voi ilmaantua yhdeksän genotyyppiluokkaa, joista syntyy neljä fenotyyppiluokkaa.
Mene luennot nro 16"Seksuaalisesti lisääntyvien monisoluisten eläinten ontogeneesi"
Mene luennot nro 18"ketjutettu perintö"
Tämä artikkeli kuvaa lyhyesti ja selkeästi Mendelin kolmea lakia. Nämä lait ovat kaiken genetiikan perusta luomalla ne, Mendel itse asiassa loi tämän tieteen.
Täältä löydät määritelmän jokaisesta laista ja opit jotain uutta genetiikasta ja biologiasta yleensä.
Ennen kuin aloitat artikkelin lukemisen, sinun tulee ymmärtää, että genotyyppi on organismin geenien kokonaisuus ja fenotyyppi sen ulkoiset ominaisuudet.
Kuka Mendel on ja mitä hän teki?
Gregor Johann Mendel on kuuluisa itävaltalainen biologi, syntynyt vuonna 1822 Gincicen kylässä. Hän opiskeli hyvin, mutta hänen perheensä oli taloudellisissa vaikeuksissa. Käsitelläkseen heitä Johann Mendel päätti vuonna 1943 ryhtyä munkkiksi tšekkiläiseen luostariin Brnon kaupungissa ja sai siellä nimen Gregor.

Gregor Johann Mendel (1822-1884)
Myöhemmin hän opiskeli biologiaa Wienin yliopistossa ja päätti sitten opettaa fysiikkaa ja luonnonhistoriaa Brnossa. Samaan aikaan tiedemies kiinnostui kasvitieteestä. Hän suoritti kokeita herneiden risteyttämisestä. Näiden kokeiden tulosten perusteella tiedemies johti kolme perinnöllisyyslakia, jotka ovat tämän artikkelin aihe.
Nämä lait, jotka julkaistiin teoksessa "Kasvihybrideillä" vuonna 1866, eivät saaneet laajaa julkisuutta, ja teos unohdettiin pian. Se muistettiin vasta Mendelin kuoleman jälkeen vuonna 1884. Tiedät jo kuinka monta lakia hän johti. Nyt on aika siirtyä tarkastelemaan jokaista.
Mendelin ensimmäinen laki - ensimmäisen sukupolven hybridien yhtenäisyyden laki
Harkitse Mendelin suorittamaa koetta. Hän otti kahdenlaisia herneitä. Nämä lajit erottuivat kukkien väristä. Toisessa ne olivat violetit ja toisessa valkoiset.
Ristitettyään ne tiedemies näki, että kaikilla jälkeläisillä oli violetit kukat. Ja keltaiset ja vihreät herneet tuottivat täysin keltaisia jälkeläisiä. Biologi toisti kokeen useita kertoja tarkistaen eri ominaisuuksien periytymisen, mutta tulos oli aina sama.

Näiden kokeiden perusteella tiedemies johti ensimmäisen lakinsa, tässä on sen muotoilu: kaikki ensimmäisen sukupolven hybridit perivät aina vain yhden ominaisuuden vanhemmiltaan.
Nimetään violeteista kukista vastaava geeni A:ksi ja valkoisista kukista a. Yhden vanhemman genotyyppi on AA (violetti) ja toisen vanhemman genotyyppi on aa (valkoinen). A-geeni peritään ensimmäiseltä vanhemmalta ja a toiselta vanhemmalta. Tämä tarkoittaa, että jälkeläisten genotyyppi on aina Aa. Isolla kirjaimella merkittyä geeniä kutsutaan dominanttiksi ja pientä kirjainta resessiiviseksi.
Jos organismin genotyyppi sisältää kaksi hallitsevaa tai kaksi resessiivistä geeniä, sitä kutsutaan homotsygoottiseksi ja eri geenejä sisältävää organismia kutsutaan heterotsygoottiseksi. Jos organismi on heterotsygoottinen, niin isolla kirjaimella merkitty resessiivinen geeni tukahdutetaan vahvemmalla dominantilla, minkä seurauksena ilmestyy se piirre, josta dominantti on vastuussa. Tämä tarkoittaa, että herneillä, joiden genotyyppi on Aa, on violetit kukat.
Kahden heterotsygoottisen organismin risteyttäminen, joilla on erilaiset ominaisuudet, on monohybridiristeytys.
Yhteisdominanssi ja epätäydellinen dominanssi
On mahdollista, että hallitseva geeni ei voi tukahduttaa resessiivistä geeniä. Ja sitten molemmat vanhempien ominaisuudet ilmestyvät kehoon.
Tämä ilmiö voidaan havaita kamelian esimerkissä. Jos tämän kasvin genotyypissä yksi geeni on vastuussa punaisista terälehdistä ja toinen valkoisista, puolet kamelian terälehdistä muuttuu punaisiksi ja loput valkoisiksi.

Tätä ilmiötä kutsutaan kodominanssiksi.
Epätäydellinen dominanssi on samanlainen ilmiö, jossa ilmenee kolmas ominaisuus, jotain vanhemmilla olleen väliltä. Esimerkiksi yökauneuskukka, jonka genotyyppi sisältää sekä valkoisia että punaisia terälehtiä, muuttuu vaaleanpunaiseksi.
Mendelin toinen laki - erottelulaki
Muistamme siis, että kun risteytetään kaksi homotsygoottista organismia, kaikki jälkeläiset saavat vain yhden piirteen. Mutta entä jos otamme kaksi heterotsygoottista organismia tästä jälkeläisestä ja risteyttäisimme ne? Ovatko jälkeläiset yhtenäiset?

Palataanpa herneisiin. Jokainen vanhempi välittää yhtä todennäköisesti joko geenin A tai geenin a. Sitten jälkeläiset jaetaan seuraavasti:
- AA - violetit kukat (25%);
- aa - valkoiset kukat (25%);
- Aa - violetit kukat (50%).
Voidaan nähdä, että purppurakukkisia organismeja on kolme kertaa enemmän. Tämä on jakautuva ilmiö. Tämä on Gregor Mendelin toinen laki: kun heterotsygoottiset organismit risteytetään, jälkeläiset jakautuvat suhteessa 3:1 fenotyypin osalta ja 1:2:1 genotyypin suhteen.
On kuitenkin olemassa niin sanottuja tappavia geenejä. Jos ne ovat läsnä, tapahtuu poikkeama toisesta laista. Esimerkiksi keltaisten hiirten jälkeläiset jaetaan suhteessa 2:1.
Sama tapahtuu platinaväristen kettujen kanssa. Tosiasia on, että jos näiden (ja joidenkin muiden) organismien genotyypissä molemmat geenit ovat hallitsevia, ne yksinkertaisesti kuolevat. Tämän seurauksena dominoiva geeni voidaan ilmentää vain, jos organismi on heterotsyoottinen.
Sukusolujen puhtauden laki ja sen sytologinen perusta
Otetaan keltaiset herneet ja vihreät herneet, keltainen geeni on hallitseva ja vihreä geeni on resessiivinen. Hybridi sisältää molemmat nämä geenit (vaikka näemme vain hallitsevan ilmentymän).

Tiedetään, että geenit siirtyvät vanhemmilta jälkeläisille sukusolujen avulla. Sukusolu on sukupuolisolu. Hybridigenotyypissä on kaksi geeniä, ja käy ilmi, että jokainen sukusolu - ja niitä on kaksi - sisälsi yhden geenin. Yhdistettyään he muodostivat hybridigenotyypin.
Jos toisessa sukupolvessa yhdelle emoorganismista tyypillinen resessiivinen ominaisuus ilmestyi, seuraavat ehdot täyttyivät:
- hybridien perinnölliset tekijät eivät muuttuneet;
- jokainen sukusolu sisälsi yhden geenin.
Toinen kohta on sukusolujen puhtauden laki. Tietenkään ei ole olemassa kahta geeniä, niitä on enemmän. On olemassa käsite alleelisista geeneistä. He ovat vastuussa samasta merkistä. Tietäen tämän käsitteen voimme muotoilla lain seuraavasti: yksi alleelista satunnaisesti valittu geeni tunkeutuu sukusoluun.
Tämän säännön sytologinen perusta: solut, joissa on kromosomeja, jotka sisältävät alleelipareja kaiken geneettisen tiedon kanssa, jakautuvat ja muodostavat soluja, joissa on vain yksi alleeli - haploidisolut. Tässä tapauksessa nämä ovat sukusoluja.
Mendelin kolmas laki - itsenäisen perinnön laki
Kolmannen lain toteutuminen on mahdollista dihybridiristeytyksellä, kun ei tutkita yhtä ominaisuutta, vaan useita. Herneiden kohdalla tämä on esimerkiksi siementen väri ja sileys.

Merkitsemme siementen väristä vastuussa olevat geenit A (keltainen) ja a (vihreä); sileyden takaamiseksi - B (sileä) ja b (ryppyinen). Yritetään tehdä dihybridiristeyttäminen organismeista, joilla on erilaisia ominaisuuksia.
Ensimmäistä lakia ei rikota tällaisen risteyttämisen aikana, eli hybridit ovat identtisiä sekä genotyypiltään (AaBb) että fenotyypiltään (keltaisilla sileillä siemenillä).
Millainen ero tulee olemaan toisessa sukupolvessa? Selvittääksesi sinun on selvitettävä, mitä sukusoluja emo-organismit voivat erittää. Ilmeisesti nämä ovat AB, Ab, aB ja ab. Tämän jälkeen rakennetaan piiri nimeltä Pinnett-hila.
Kaikki sukusolut, jotka yksi organismi voi vapauttaa, on lueteltu vaakasuunnassa, ja kaikki sukusolut, jotka toinen organismi voi vapauttaa, on listattu pystysuoraan. Verkon sisällä kirjataan organismin genotyyppi, joka esiintyisi annettujen sukusolujen kanssa.
| AB | Ab | aB | ab | |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | AAbb | AaBb | Aabb |
| aB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
Jos tutkit taulukkoa, voit päätellä, että toisen sukupolven hybridien jakautuminen fenotyypin mukaan tapahtuu suhteessa 9:3:3:1. Mendel ymmärsi tämän myös useiden kokeiden jälkeen.
Lisäksi hän tuli myös siihen tulokseen, että se, mikä yhden alleelin (Aa) geeneistä pääsee sukusoluun, ei riipu toisesta alleelista (Bb), eli on olemassa vain itsenäinen ominaisuuksien periytyminen. Tämä on hänen kolmas lakinsa, jota kutsutaan itsenäisen perinnön laiksi.
Johtopäätös
Mendelin kolme lakia ovat geneettisiä peruslakeja. Kiitos siitä, että yksi henkilö päätti kokeilla herneitä, biologia sai uuden osan - genetiikka.
Sen avulla tiedemiehet kaikkialta maailmasta ovat oppineet monia asioita sairauksien ehkäisystä geenitekniikkaan. Genetiikka on yksi mielenkiintoisimmista ja lupaavimmista biologian aloista.
Mendelin kolmas laki on ominaisuuksien riippumattoman jakautumisen laki. Tämä tarkoittaa, että jokainen yhden alleeliparin geeni voi esiintyä sukusolussa minkä tahansa muun geenin kanssa toisesta alleeliparista. Esimerkiksi, jos organismi on heterotsygoottinen kahdelle tutkittavalle geenille (AaBb), se muodostaa seuraavan tyyppisiä sukusoluja: AB, Ab, aB, ab. Toisin sanoen esimerkiksi geeni A voi olla samassa sukusolussa sekä geenin B:n että b:n kanssa. Sama koskee muita geenejä (niiden mielivaltainen yhdistelmä ei-alleelisten geenien kanssa).
Mendelin kolmas laki on jo ilmeinen dihybridiristeyksellä(etenkin trihybridin ja polyhybridin kanssa), kun puhtaat linjat eroavat kahdesta tutkitusta ominaisuudesta. Mendel risteytti hernelajikkeen keltaisilla sileillä siemenillä lajikkeella, jossa oli vihreitä ryppyisiä siemeniä ja josta saatiin yksinomaan keltaisia sileitä siemeniä F 1 . Seuraavaksi hän kasvatti siemenistä F 1 -kasveja, antoi niiden itsepölytyttää ja hankki F 2 -siemeniä. Ja täällä hän havaitsi halkeilua: kasveissa oli sekä vihreitä että ryppyisiä siemeniä. Yllättävintä oli, että toisen sukupolven hybridien joukossa ei ollut vain kasveja, joilla oli sileät keltaiset ja vihreät ryppyiset siemenet. Siellä oli myös keltaisia ryppyisiä ja vihreitä sileitä siemeniä, eli tapahtui merkkien rekombinaatioita ja saatiin yhdistelmiä, joita ei löytynyt alkuperäisistä vanhempainmuodoista.
Analysoimalla eri F2-siementen kvantitatiivista suhdetta Mendel havaitsi seuraavan:
Jos tarkastellaan jokaista ominaisuutta erikseen, se jaettiin suhteessa 3:1, kuten monohybridiristeyksessä. Eli jokaista kolmea keltaista siementä kohden oli yksi vihreä ja jokaista 3 sileää siementä kohti yksi ryppyinen.
Kasveja, joilla oli uusia ominaisuuksia, ilmestyi.
Fenotyyppisuhde oli 9:3:3:1, jossa jokaista yhdeksää keltaista sileää herneensiementä kohden oli kolme keltaista ryppyistä, kolme vihreää sileää ja yksi vihreä ryppyinen.
Mendelin kolmatta lakia havainnollistaa hyvin Punnettin hila. Täällä vanhempien mahdolliset sukusolut (tässä tapauksessa ensimmäisen sukupolven hybridit) kirjoitetaan rivi- ja sarakeotsikoihin. Kunkin sukusolutyypin tuottamisen todennäköisyys on ¼. On myös yhtä todennäköistä, että ne yhdistyvät eri tavalla yhdeksi tsygootiksi.
Näemme, että muodostuu neljä fenotyyppiä, joista kahta ei ollut aiemmin olemassa. Fenotyyppien suhde on 9:3:3:1. Eri genotyyppien lukumäärä ja niiden suhde on monimutkaisempi:
Tämä johtaa 9 eri genotyyppiin. Niiden suhde on: 4:2:2:2:2:1:1:1:1. Samaan aikaan heterotsygootit ovat yleisempiä ja homotsygootit harvinaisempia.
Jos palataan siihen tosiasiaan, että jokainen ominaisuus periytyy itsenäisesti ja jokaiselle havaitaan 3:1-jako, niin voimme laskea fenotyyppien todennäköisyyden kahdelle eri alleelin ominaisuudelle kertomalla kunkin alleelin ilmenemistodennäköisyys (ts. Punnettin hilaa ei tarvitse käyttää). Siten tasaisten keltaisten siementen todennäköisyys on ¾ × ¾ = 9/16, sileiden vihreiden - ¾ × ¼ = 3/16, ryppyjen keltaisten - ¼ × ¾ = 3/16, ryppyjen vihreiden - ¼ × ¼ = 1 /16. Siten saamme saman fenotyyppisuhteen: 9:3:3:1.
Mendelin kolmas laki selittyy eri parien homologisten kromosomien itsenäisellä erolla meioosin ensimmäisen jakautumisen aikana. Kromosomi, joka sisältää geenin A, voi yhtä suurella todennäköisyydellä mennä samaan soluun sekä kromosomin sisältävän geenin B että kromosomin sisältävän geenin b kanssa. Kromosomi, jossa on geeni A, ei ole millään tavalla liitetty geenin B kromosomiin, vaikka ne molemmat periytyivät samalta vanhemmalta. Voimme sanoa, että meioosin seurauksena kromosomit sekoittuvat. Niiden erilaisten yhdistelmien lukumäärä lasketaan kaavalla 2 n, jossa n on haploidisen joukon kromosomien lukumäärä. Joten jos lajilla on kolme paria kromosomeja, niin niiden erilaisten yhdistelmien lukumäärä on 8 (2 3).
Kun ominaisuuksien itsenäisen periytymisen laki ei päde
Mendelin kolmas laki eli ominaisuuksien itsenäisen periytymisen laki koskee vain geenejä, jotka sijaitsevat eri kromosomeissa tai sijaitsevat samassa kromosomissa, mutta melko kaukana toisistaan.
Periaatteessa, jos geenit sijaitsevat samassa kromosomissa, ne periytyvät yhdessä, eli niillä on yhteys toisiinsa, eikä ominaisuuksien itsenäisen periytymisen laki enää päde.
Esimerkiksi jos herneensiementen väristä ja muodosta vastaavat geenit olisivat samassa kromosomissa, niin ensimmäisen sukupolven hybridit saattoivat muodostaa vain kahden tyyppisiä sukusoluja (AB ja ab), koska meioosin aikana vanhemman kromosomit eroavat toisistaan riippumatta. , mutta ei yksittäisiä geenejä. Tässä tapauksessa toisessa sukupolvessa jakautuminen olisi 3:1 (kolme keltaista sileää yhteen vihreään ryppyiseen).
Se ei kuitenkaan ole niin yksinkertaista. Koska kromosomien konjugaatio (yhteen saattaminen) ja risteytys (kromosomiosien vaihto) esiintyy luonnossa, myös homologisissa kromosomeissa sijaitsevat geenit yhdistyvät uudelleen. Joten jos kromosomi, jossa on AB-geenejä, vaihtaa risteytysprosessin aikana osan B-geenin kanssa homologiseen kromosomiin, jonka osa sisältää b-geenin, voidaan uusia sukusoluja (Ab ja esimerkiksi aB). saatu. Tällaisten rekombinanttisukusolujen prosenttiosuus on pienempi kuin jos geenit olisivat eri kromosomeissa. Tässä tapauksessa ylityksen todennäköisyys riippuu kromosomissa olevien geenien etäisyydestä: mitä kauempana, sitä suurempi on todennäköisyys.
1. Kuvaile G. Mendelin ensimmäinen ja toinen laki.
Mendelin ensimmäinen laki (ensimmäisen sukupolven hybridien yhtenäisyyslaki) sanoo, että kun risteytetään alleelisten ominaisuuksien suhteen erilaisia organismeja, vain yksi niistä ilmestyy ensimmäisen sukupolven hybrideihin - hallitseva, kun taas vaihtoehto, resessiivinen, pysyy piilossa.
Mendelin toinen laki (segregaatiolaki) sanoo, että kun kaksi ensimmäisen sukupolven hybridiä risteytetään keskenään, molemmat alkuperäisen vanhempainmuodon ominaisuudet näkyvät tietyssä suhteessa toisessa sukupolvessa.
2. Mitä yhtäläisyyksiä ja eroja G. Mendelin kolmannella lailla ja T. Morganin lailla on?
Samankaltaisuus piilee siinä, että sekä Mendelin kolmas laki että Morganin laki käsittelevät kahden erilaisen ominaisuuden periytymistä. Erot ovat siinä, että Mendelin laissa kyseessä on itsenäinen perinnöllisyys, kun taas Morganin laissa puhutaan ominaisuuksien linkitetystä (yhteisestä) periytymisestä.
3. Mikä on perinnöllisyyden ja vaihtelevuuden rooli elävässä luonnossa?
Perinnöllisyyden ansiosta lajien säilyminen varmistetaan merkittävinä ajanjaksoina (jopa satoja miljoonia vuosia). Ympäristöolosuhteet kuitenkin muuttuvat (joskus merkittävästi) ajan myötä, ja tällaisissa tapauksissa lajin yksilöiden monimuotoisuuteen johtava vaihtelu varmistaa lajin selviytymisen. Jotkut yksilöistä osoittautuvat paremmin sopeutuneiksi uusiin olosuhteisiin, ja tämä antaa heille mahdollisuuden selviytyä. Lisäksi vaihtelevuuden ansiosta lajit voivat laajentaa elinympäristönsä rajoja ja kehittää uusia alueita.
4. Ovatko G. Mendelin lait universaaleja ja soveltuvatko ne ihmisiin?
Joo. Mendelin lait ovat yleismaailmallisia ja pätevät kaikkiin eläviin organismeihin.
5. Kuvaile käsitteiden "geeni", "alleeli", "risteytys" välistä suhdetta.
Geenit ovat DNA-molekyylin osa, joka vastaa organismin yhden proteiinin tai RNA:n rakentamisesta. Alleelit ovat saman geenin tilan muotoja, jotka miehittävät identtisiä lokuksia homologisissa kromosomeissa ja aiheuttavat fenotyyppisiä eroja samassa ominaisuudessa. Crossing over on homologisten kromosomien homologisten osien risteytys, keskinäinen vaihto.
6. Mikä on mutaatio? Milloin ja missä mutaatioita tapahtuu?
Mutaatiot ovat äkillisiä periytyviä muutoksia geneettisessä materiaalissa, jotka aiheuttavat muutoksia organismin ominaisuuksissa ja ominaisuuksissa. Mutaatiot voivat olla luonnollisia, spontaaneja, eli tapahtuvia tahattomasti tai keinotekoisia, jotka tapahtuvat, kun organismit altistuvat erilaisille tekijöille - mutageeneille.
7. Millaisia ylityksiä G. Mendel tutki?
Monohybridi- ja dihybridiristeys.
8. Mikä on geenien moninkertaisen toiminnan erikoisuus?
Pleiotropia on useiden geenitoimintojen ilmiö. Se ilmenee yhden geenin kyvyssä vaikuttaa useisiin fenotyyppisiin piirteisiin. Esimerkki: geeni, joka vastaa punaisen pigmentin muodostumisesta kukassa, edistää sen ilmestymistä varteen ja lehtiin, aiheuttaa varren pidentymistä ja siementen massan kasvua.
 Verbin aikamuodot eivät muutu henkilökohtaisesti
Verbin aikamuodot eivät muutu henkilökohtaisesti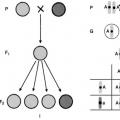 Mendelin lakien täyttymisen ehdot
Mendelin lakien täyttymisen ehdot Jakava pehmeä merkki
Jakava pehmeä merkki