Mendelin jakamissääntö. Genetiikan perusteet: Mendelin lait. Gregor Johann Mendelin elämä ja tieteellinen tutkimus
Mendelin ensimmäinen laki
Kahden organismin risteytymistä kutsutaan hybridisaatio, jälkeläisiä, jotka ovat peräisin kahden eri perinnöllisen yksilön risteyttämisestä, kutsutaan hybridi, ja erillinen henkilö - hybridi. Mono hybridi kutsutaan kahden organismin risteytykseksi, jotka eroavat toisistaan yhdellä vaihtoehtoisten (toisensa poissulkevien) ominaisuuksien parilla. Näin ollen tällaisella risteyttämisellä voidaan jäljittää vain kahden ominaisuuden periytymismalleja, joiden kehittymisen määrää alleelisten geenien pari. Kaikkia muita näille organismeille ominaisia ominaisuuksia ei oteta huomioon.
Jos ristät hernekasvit keltaisilla ja vihreillä siemenillä, kaikilla tuloksena olevilla hybrideillä on keltaisia siemeniä. Sama kuva havaitaan risteyttäessä kasveja sileillä ja ryppyisillä siemenillä; kaikilla ensimmäisen sukupolven jälkeläisillä on tasainen siemenmuoto:
Siksi Mendelin ensimmäinen laki kutsuttiin Ensimmäisen sukupolven hybridien yhtenäisyyslaki.
Jos käytetään termejä, jotka ilmestyivät monta vuotta Mendelin työn jälkeen, voidaan sanoa, että yhden lajikkeen hernekasvien solut sisältävät kaksi geeniä vain keltaiseen värjäykseen ja toisen lajikkeen kasvien geenit sisältävät kaksi geeniä vain vihreään värjäämiseen. Genejä, jotka vastaavat yhden ominaisuuden (esimerkiksi siemenvärin) kehittymisestä, kutsutaan alleeliset geenit. Näin ollen ensimmäisen sukupolven hybridissä vain yksi kustakin vaihtoehtoisten hahmojen parista kehittyy. Toinen merkki näyttää katoavan eikä näy. Valta-ilmiö hybridissä klo toisen vanhemman merkki G. Mendel nimeltään Dominie kiertävä. Ominaisuutta, joka esiintyy ensimmäisen sukupolven hybridissä ja estää toisen ominaisuuden kehittymisen, kutsuttiin hallitseva, ja päinvastainen eli tukahdutettu merkki - resessiivinen. Gregor Mendel päätti korvata kasvin ominaisuuksien kuvauksen abstraktilla koodilla "A, B, C, D, E, F, G" ja "a, b, c, d, e, f, g" ja sitten huomioimatta yhden hahmoparin kohtalo hän siirtyi tarkkailemaan kahta, kolmea, neljää paria samanaikaisesti. Suurilla A, B, C, D, E, F, G hän nimesi hallitsevia ominaisuuksia; pienet a, b, c, d, e, f, g ovat resessiivisiä. Jos organismin genotyypissä (tsygootissa) on kaksi identtistä alleelista geeniä - molemmat hallitsevia tai molemmat resessiivisiä (AA tai aa), sellaista organismia kutsutaan homotsygoottinen. Jos alleelisten geenien parista toinen on hallitseva ja toinen on resessiivinen (Ah) silloin sellaista organismia kutsutaan heterotsygoottinen.
Erottelulaki tai Mendelin toinen laki.
Jos ensimmäisen sukupolven jälkeläiset, jotka ovat identtisiä tutkittavassa ominaisuudessa, risteytetään keskenään, niin toisessa sukupolvessa molempien vanhempien ominaisuudet esiintyvät tietyssä numerosuhteessa: 3/4 yksilöistä on hallitseva piirre, ¼ resessiivinen:

Ilmiö, jossa risteytys heterotsygoottinen Yksilöt johtavat jälkeläisten muodostumiseen, joista osa sarvella on hallitseva ominaisuus ja osa - resessiivinen ny, nimeltään splitting. Tämän seurauksena resessiivinen ominaisuus ei hävinnyt ensimmäisen sukupolven hybrideissä, vaan se vain tukahdutettiin ja tulee esiin toisessa hybridisukupolvessa.
Hypoteesi sukusolujen puhtaudesta. Mendel ehdotti, että hybridien muodostumisen aikana perinnölliset tekijät eivät sekoitu, vaan pysyvät ennallaan. Hybridissä molemmat tekijät ovat läsnä - hallitseva ja resessiivinen, mutta hallitseva perinnöllinen tekijä ilmenee piirteenä, kun taas resessiivinen on tukahdutettu. Sukupolvien välinen viestintä seksuaalisen lisääntymisen aikana tapahtuu sukusolujen kautta - ha meta. Siksi on oletettava, että jokainen sukusolu kantaa vain yhden tekijän parista. Sitten hedelmöityksen aikana kahden sukusolun fuusio, joista kummassakin on resessiivinen perinnöllinen tekijä, johtaa organismin muodostumiseen, jolla on fenotyyppisesti ilmenevä resessiivinen piirre. Sukusolujen fuusio, joista jokaisessa on hallitseva tekijä, tai kaksi sukusolua, joista toinen sisältää hallitsevan ja toinen resessiivisen tekijän, johtaa organismin kehittymiseen, jolla on hallitseva piirre.
Mendel selitti jälkeläisten jakautumisen heterotsygoottisten yksilöiden risteyttämisessä sillä, että sukusolut ovat geneettisesti puhtaita, eli ne sisältävät vain yhden geenin alleeliparista. hypoteesi(nykyään laki) puhtaus sukusolut voidaan formuloida seuraavasti: sukusolujen muodostumisen aikana vain yksi geeni alleeliparista tulee jokaiseen sukusoluun.
Miksi ja miten tämä tapahtuu? Tiedetään, että jokaisessa kehon solussa on täsmälleen sama diploidisarja kromosomeja. Kaksi homologista kromosomia sisältää kaksi identtistä geeniä. Geneettisesti "puhtaat" sukusolut muodostuvat seuraavasti: kun uros- ja naaraspuoliset sukusolut yhdistyvät, saadaan hybridi, jossa on diploidi (kaksois) kromosomisarja.
Kuten kaaviosta (Liite 2) voidaan nähdä, tsygootti saa puolet kromosomeista isän kehosta ja puolet äidin kehosta.
Sukusolujen muodostumisen aikana hybridissä myös homologiset kromosomit päätyvät eri soluihin ensimmäisen meioottisen jakautumisen aikana.
Tämän alleeliparin perusteella muodostuu kahden tyyppisiä sukusoluja. Hedelmöityksen aikana sukusolut, joissa on sama tai eri alleeli, kohtaavat toisensa sattumalta. Tilastollisen todennäköisyyden vuoksi riittävän suurella määrällä sukusoluja jälkeläisissä genotyypeistä 25 % on homotsygoottisia dominansseja, 50 % heterotsygoottisia, 25 % homotsygoottisia resessiivisiä, eli suhde on vakiintunut. 1AA:2Aa:1aa.
Vastaavasti, fenotyypin mukaan, toisen sukupolven jälkeläiset monohybridiristeytyksen aikana jakautuvat suhteessa 3:1 (¾ yksilöä, joilla on hallitseva piirre, ¼ yksilöä, joilla on resessiivinen piirre).
Sytologinen perusta merkkien erottelulle monohybridiristeytyksen aikana on homologisten kromosomien hajoaminen solun eri napoihin ja haploidisten sukusolujen muodostuminen meioosissa.
Yllä käsitellyissä esimerkeissä yhtenäisyyssääntö ilmaistiin siinä, että kaikki hybridit olivat ulkonäöltään samanlaisia kuin yksi vanhemmista. Tätä ei aina huomioida. Usein heterotsygoottisten muotojen ominaisuudet ovat luonteeltaan keskitasoisia, ts. määräävä asema ei välttämättä ole täydellinen. Kaavio Night Beauty -kasvin kahden perinnöllisen muodon risteyttämisestä:


Yhdessä niistä on punaisia kukkia (ja tämä on hallitseva ominaisuus), ja toisessa on valkoisia kukkia. Kaavio osoittaa, että kaikilla ensimmäisen sukupolven hybrideillä on vaaleanpunaiset kukat. Toisessa sukupolvessa halkeaminen tapahtuu suhteessa 1:2:1, ts. yksi punainen kukka (homotsygootti), kaksi vaaleanpunaista kukkaa (heterotsygootti), yksi valkoinen (homotsygootti). Tätä ilmiötä kutsutaan epätäydelliseksi dominanssiksi.
Epätäydellisen dominanssin tapauksessa heterotsygoottisessa tilassa hallitseva geeni ei aina täysin suppressoi resessiivistä geeniä. Joissakin tapauksissa hybridi fi ei toista täysin mitään vanhemmuuden ominaisuuksista ja ominaisuus on luonteeltaan keskitasoinen, ja se poikkeaa enemmän tai vähemmän kohti hallitsevaa tai resessiivistä tilaa. Mutta kaikki tämän sukupolven yksilöt ovat yhtenäisiä tässä ominaisuudessa. Epätäydellinen dominointi on laajalle levinnyt ilmiö. Se havaittiin tutkittaessa kukkavärin perinnöllisyyttä lohikäärmeillä, villan värin periytymistä nautaeläimillä ja lampailla, biokemiallisia ominaisuuksia ihmisillä jne. Epätäydellisen dominanssin seurauksena ilmenevät väliominaisuudet edustavat usein esteettistä tai aineellista arvoa ihmisille. Herää kysymys: onko valinnalla mahdollista kehittää esimerkiksi erilaisia yökauneutta vaaleanpunaisella kukkavärillä? Ilmeisesti ei, koska tämä ominaisuus kehittyy vain heterotsygooteissa ja kun ne risteytetään keskenään, halkeilu tapahtuu aina:
Epätäydellinen dominanssi on laajalle levinnyt ilmiö. Se havaittiin tutkittaessa kukkavärin perinnöllisyyttä lohikäärmeillä, villan värin periytymistä nautaeläimillä ja lampailla, biokemiallisia ominaisuuksia ihmisillä jne. Epätäydellisen dominanssin seurauksena ilmenevät väliominaisuudet edustavat usein esteettistä tai aineellista arvoa ihmisille. Herää kysymys: onko valinnalla mahdollista kehittää esimerkiksi erilaisia yökauneutta vaaleanpunaisella kukkavärillä? Ilmeisesti ei, koska tämä ominaisuus kehittyy vain heterotsygooteissa ja kun ne risteytetään keskenään, halkeilu tapahtuu aina.
Itsenäisen yhdistelyn laki tai kolmas Mendelin laki.
Mendelin tutkimus yhden alleeliparin periytymisestä mahdollisti joukon tärkeitä geneettisiä malleja: dominanssiilmiön, resessiivisten alleelien pysyvyyden hybrideissä, hybridien jälkeläisten jakautumisen suhteessa 3:1, ja myös olettaa, että sukusolut ovat geneettisesti puhtaita, eli ne sisältävät vain yhden geenin alleelipareista. Organismit eroavat kuitenkin monista geeneistä. On mahdollista luoda kahden tai useamman vaihtoehtoisen merkkiparin periytymismalleja dihybridi tai polyhybridiristeys, ts. vanhempien muotojen risteytys, jotka eroavat kahdella ominaisuusparilla.
Gregor Johann Mendel(1822-1884) - itävaltalainen tiedemies, genetiikan perustaja. Hän käytti ensimmäistä kertaa hybridologista menetelmää ja havaitsi perinnöllisten tekijöiden olemassaolon, joita myöhemmin kutsutaan geeneiksi.
Mendelin ensimmäinen laki
G. Mendel risteytti hernekasveja keltaisilla siemenillä ja hernekasveja vihreillä siemenillä. Molemmat olivat puhtaita linjoja, ts. homotsygoottinen:
Mendelin ensimmäinen laki on ensimmäisen sukupolven hybridien yhtenäisyyden laki (dominanssilaki): kun puhtaat linjat risteytyvät, kaikilla ensimmäisen sukupolven hybrideillä on yksi ominaisuus (dominoiva).
Mendelin toinen laki
Tämän jälkeen G. Mendel risteytti ensimmäisen sukupolven hybridit keskenään:
Mendelin toinen laki on luonteen jakautumisen laki: ensimmäisen sukupolven hybridit, kun ne risteytyvät, jaetaan tietyssä numeerisessa suhteessa: yksilöt, joilla on resessiivinen ilmentymä piirre, muodostavat ¼ jälkeläisten kokonaismäärästä.
Ilmiö, jossa heterotsygoottisten yksilöiden risteytys johtaa jälkeläisten muodostumiseen, joista osalla on hallitseva piirre ja osa on resessiivisiä, on ns. jakaminen. Kun
Monohybridiristeyksessä tämä suhde on seuraava: 1AA:2Aa:1aa, so. 3:1 (jos vallitsee täydellinen) tai 1:2:1 (jos dominanssi on epätäydellinen); dihybridiristeykseen - 9:3:3:1 tai (3:1) 2; polyhybridillä - (3:1) n.
Dominoiva geeni ei aina täysin suppressoi resessiivistä geeniä. Tätä ilmiötä kutsutaan epätäydellinen ylivalta. Esimerkki epätäydellisestä dominanssista on yökauneuskasvin kukkien värin periytyminen:
Ensimmäisen sukupolven yhtenäisyyden ja toisen sukupolven hahmojen jakautumisen sytologinen perusta
Ne koostuvat homologisten kromosomien hajoamisesta ja haploidisten sukusolujen muodostumisesta meioosissa.
Hypoteesi (laki) sukusolujen puhtaudesta
Se on seuraava:
- Kun sukusoluja muodostuu, jokaiseen sukusoluun tulee vain yksi alleeli alleeliparista, ts. sukusolut ovat geneettisesti puhtaita;
- Hybridiorganismissa geenit eivät hybridisoidu (eivät sekoitu) ja ovat puhtaassa alleelisessa tilassa.
Splitting-ilmiön tilastollinen luonne
Sukusolujen puhtauden hypoteesista seuraa, että erottelulaki on seurausta eri geenejä sisältävien sukusolujen satunnaisesta yhdistelmästä. Kun otetaan huomioon sukusolujen yhteyden satunnainen luonne, kokonaistulos osoittautuu luonnolliseksi. Tästä seuraa, että monohybridiristeyksessä suhdetta 3:1 (jos kyseessä on täydellinen dominanssi) tai 1:2:1 (jos vallitsee epätäydellinen dominanssi) on pidettävä tilastollisiin ilmiöihin perustuvana mallina. Tämä pätee myös polyhybridiristeykseen. Numeeristen suhteiden tarkka toteutus halkaisun aikana on mahdollista vain suurella määrällä tutkittavia hybridiyksilöitä. Siten genetiikan lait ovat luonteeltaan tilastollisia.
Jälkeläisten analyysi
Analyysi risti avulla voit määrittää, onko organismi homotsygoottinen vai heterotsygoottinen hallitsevan geenin suhteen. Tätä varten yksilö, jonka genotyyppi on määritettävä, risteytetään resessiivisen geenin homotsygoottisen yksilön kanssa. Usein toinen vanhemmista risteytetään toisen jälkeläisen kanssa. Tätä risteystä kutsutaan palautettavissa.
Dominoivan yksilön homotsygoottisuuden tapauksessa jakautumista ei tapahdu:
Mendelin kolmas laki
G. Mendel suoritti dihybridiristeyttämisen hernekasveista, joissa oli keltaisia ja sileitä siemeniä, ja hernekasveja, joissa oli vihreitä ja ryppyisiä siemeniä (molemmat puhtaat linjat), ja sitten risteytti niiden jälkeläiset. Tämän seurauksena hän havaitsi, että jokainen hahmopari käyttäytyy jälkeläisiksi jaettuna samalla tavalla kuin monohybridiristeyksessä (jakaantuu 3:1), ts. riippumatta toisesta merkkiparista.
Tuloksena saatiin seuraavat tiedot:
- 9/16 - hernekasvit keltaisilla sileillä siemenillä;
- 3/16 - hernekasvit keltaisilla ryppyisillä siemenillä;
- 3/16 - hernekasvit vihreillä sileillä siemenillä;
- 1/16 - hernekasvit vihreillä ryppyisillä siemenillä.
Mendelin kolmas laki on ominaisuuksien itsenäisen yhdistelmän (perinnön) laki: kunkin ominaisuuden erottelu tapahtuu muista ominaisuuksista riippumatta.
Riippumattoman yhdistelmän sytologinen perusta on kunkin parin homologisten kromosomien poikkeamisen satunnainen luonne solun eri napoihin meioosin aikana, riippumatta muista homologisten kromosomien pareista. Tämä laki pätee vain, jos erilaisten ominaisuuksien kehittymisestä vastaavat geenit sijaitsevat eri kromosomeissa. Poikkeuksia ovat linkitetyt perinnöt.
Tšekkiläinen tutkimusmatkailija Gregor Mendel(1822-1884) harkitaan genetiikan perustaja, koska hän oli ensimmäinen, jo ennen tämän tieteen muotoutumista, joka muotoili perinnön peruslait. Monet tiedemiehet ennen Mendeliä, mukaan lukien erinomainen saksalainen 1700-luvun hybridisaattori. I. Kelreuter totesi, että eri lajikkeisiin kuuluvia kasveja risteyttäessä hybridijälkeläisissä havaitaan suurta vaihtelua. Kukaan ei kuitenkaan pystynyt selittämään monimutkaista jakamista ja lisäksi pelkistämään sitä tarkkoihin kaavoihin, koska hybridologisen analyysin tieteellistä menetelmää ei ollut.
Hybridologisen menetelmän kehityksen ansiosta Mendel onnistui välttämään aiempia tutkijoita hämmentäneitä vaikeuksia. G. Mendel raportoi työnsä tuloksista vuonna 1865 luonnontieteilijöiden seuran kokouksessa Brünnissä. Itse teos, nimeltään "Experiments on Plant Hybrids", julkaistiin myöhemmin tämän yhteiskunnan "Proceedings" -lehdessä, mutta se ei saanut asianmukaista arviointia aikalaisilta ja jäi unohduksiin 35 vuodeksi.
Munkina G. Mendel suoritti klassisia kokeitaan eri hernelajikkeiden risteyttämisestä Brünnin luostarin puutarhassa. Hän valitsi 22 hernelajiketta, joilla oli selkeät vaihtoehtoiset erot seitsemässä ominaisuudessa: siemenet keltaiset ja vihreät, sileät ja ryppyiset, kukat punaiset ja valkoiset, kasvit korkeat ja lyhyet jne. Hybridologisen menetelmän tärkeä ehto oli puhtaan eli vanhempana pakollinen käyttö. muotoja, jotka eivät hajoa tutkittujen ominaisuuksien mukaan.
Onnistuneella kohteen valinnalla oli suuri rooli Mendelin tutkimuksen onnistumisessa. Herneet ovat itsepölyttäjiä. Ensimmäisen sukupolven hybridien saamiseksi Mendel kastroi emokasvin kukat (poisti ponnet) ja pölytti emit keinotekoisesti urosvanhemman siitepölyllä. Kun hankittiin toisen sukupolven hybridejä, tämä menettely ei ollut enää tarpeen: hän yksinkertaisesti jätti F 1 -hybridit itsepölyttämään, mikä teki kokeesta vähemmän työvoimavaltaisen. Hernekasvit lisääntyivät yksinomaan seksuaalisesti, joten mitkään poikkeamat eivät saaneet vääristää kokeen tuloksia. Ja lopuksi, herneistä Mendel löysi riittävän määrän kirkkaasti vastakkaisia (vaihtoehtoisia) ja helposti erotettavissa olevia hahmopareja analysoitavaksi.
Mendel aloitti analyysinsä yksinkertaisimmalla risteytystyypillä - monohybridillä, jossa vanhemmat yksilöt eroavat yhden piirreparin osalta. Ensimmäinen Mendelin löytämä periytymismalli oli, että kaikilla ensimmäisen sukupolven hybrideillä oli sama fenotyyppi ja ne perivät yhden vanhemman piirteen. Mendel kutsui tätä ominaisuutta hallitsevaksi. Toisen vanhemman vaihtoehtoista ominaisuutta, jota ei esiintynyt hybrideissä, kutsuttiin resessiiviseksi. Löydety malli nimettiin Mendelin lain I eli ensimmäisen sukupolven hybridien yhtenäisyyden laki. Toisen sukupolven analyysin aikana muodostettiin toinen malli: hybridien jakautuminen kahteen fenotyyppiseen luokkaan (dominoivalla ominaisuudella ja resessiivisellä ominaisuudella) tietyissä numeerisissa suhteissa. Laskemalla yksilöiden lukumäärän kussakin fenotyyppiluokassa Mendel totesi, että jakautuminen monohybridiristeyksessä vastaa kaavaa 3:1 (kolme kasvia, joilla on hallitseva ominaisuus, yksi resessiivinen ominaisuus). Tätä mallia kutsutaan Mendelin II laki tai erottelulaki. Kaikkien seitsemän ominaisuusparin analyysissä syntyi avoimia kaavoja, joiden perusteella kirjoittaja tuli johtopäätökseen niiden universaalisuudesta. Itsepölyttäessään F 2 -hybridejä Mendel sai seuraavat tulokset. Valkoiset kasvit tuottivat jälkeläisiä, joilla oli vain valkoisia kukkia. Punakukkaiset kasvit käyttäytyivät eri tavalla. Vain kolmasosa heistä antoi yhtenäisiä jälkeläisiä punaisilla kukilla. Loput jälkeläiset jaettiin punaisen ja valkoisen värin suhteen suhteessa 3:1.
Alla on kaavio herneenkukan värin periytymisestä, havainnollistaen Mendelin I ja II lakeja.
Yrittääkseen selittää avoimien kuvioiden sytologista perustaa Mendel muotoili ajatuksen sukusolujen sisältämistä erillisistä perinnöllisistä taipumuksista ja parillisten vaihtoehtoisten hahmojen kehityksen määrittämisestä. Jokaisessa sukusolussa on yksi perinnöllinen talletus, ts. on "puhdas". Hedelmöityksen jälkeen tsygootti saa kaksi perinnöllistä kerrostumaa (toinen äidiltä, toinen isältä), jotka eivät sekoitu ja myöhemmin, kun hybridi muodostaa sukusoluja, ne päätyvät myös eri sukusoluihin. Tätä Mendelin hypoteesia kutsuttiin "sukusolujen puhtauden säännöksi". Perinnöllisten taipumusten yhdistelmä tsygootissa määrittää, minkä luonteen hybridillä on. Mendel merkitsi isolla kirjaimella taipumusta, joka määrää hallitsevan piirteen kehittymisen ( A), ja resessiivinen kirjataan isolla ( A). Yhdistelmä AA Ja Ahh tsygootissa määrittää hallitsevan piirteen kehittymisen hybridissä. Resessiivinen ominaisuus ilmenee vain yhdistettynä ahh.
Vuonna 1902 V. Betson ehdotti, että parihahmojen ilmiö nimettäisiin termillä "allelomorfismi" ja itse hahmot vastaavasti "allelomorfisiksi". Hänen ehdotuksensa mukaan organismeja, joilla oli samat perinnölliset taipumukset, alettiin kutsua homotsygoottisiksi ja niitä, jotka sisältävät erilaisia taipumuksia - heterotsygoottisiksi. Myöhemmin termi "allelomorfismi" korvattiin lyhyemmällä termillä "allelismi" (Johansen, 1926), ja vaihtoehtoisten piirteiden kehittymisestä vastuussa olevia perinnöllisiä taipumuksia (geenejä) kutsuttiin "alleelisiksi".
Hybridologiseen analyysiin kuuluu vanhempien muotojen vastavuoroinen risteyttäminen, ts. käyttämällä samaa yksilöä ensin äidin vanhempana (eteenpäin risteytys) ja sitten isän vanhempana (backcrossing). Jos molemmat risteytykset tuottavat samat tulokset, jotka vastaavat Mendelin lakeja, tämä osoittaa, että analysoitu piirre on autosomaalinen geeni. Muuten ominaisuus liittyy sukupuoleen, koska geeni on lokalisoitunut sukupuolikromosomiin.

Kirjainmerkinnät: P - vanhempi yksilö, F - hybridi yksilö, ♀ ja ♂ - naaras tai uros yksilö (tai sukusolu),
iso kirjain (A) on hallitseva perinnöllinen taipumus (geeni), pieni kirjain (a) on resessiivinen geeni.
Toisen sukupolven hybridien joukossa, joilla on keltainen siemenväri, on sekä hallitsevia homotsygootteja että heterotsygootteja. Hybridin spesifisen genotyypin määrittämiseksi Mendel ehdotti hybridin risteyttämistä homotsygoottisen resessiivisen muodon kanssa. Sitä kutsutaan analysoimiseksi. Kun ylitetään heterotsygootti ( Ahh) analysaattorilinjalla (aa) havaitaan jakautuminen sekä genotyypin että fenotyypin mukaan suhteessa 1:1.

Jos toinen vanhemmista on homotsygoottinen resessiivinen muoto, niin analysoivasta risteyksestä tulee samanaikaisesti takaisinristeytys - hybridin paluuristeys emomuodon kanssa. Tällaisen ristin jälkeläiset on nimetty Fb.
Mendelin monohybridisten risteytysten analysoinnissa löytämät kuviot ilmenivät myös dihybridiristeyksissä, joissa vanhemmat erosivat kahdessa vaihtoehtoisessa ominaisuusparissa (esimerkiksi keltainen ja vihreä siemenväri, sileä ja ryppyinen muoto). Fenotyyppiluokkien määrä F 2:ssa kuitenkin kaksinkertaistui ja fenotyyppisen jakautumiskaava oli 9:3:3:1 (9 yksilölle, joilla oli kaksi hallitsevaa ominaisuutta, kolme yksilöä, joilla kullakin on yksi dominantti ja yksi resessiivinen ominaisuus, ja yksi yksilö, jolla on kaksi resessiiviset ominaisuudet).
F 2:n halkeamisen analysoinnin helpottamiseksi englantilainen geneetikko R. Punnett ehdotti sen graafista esitystä hilan muodossa, jota alettiin kutsua hänen nimensä mukaan ( Punett-verkko). Vasemmalla, pystysuunnassa, se sisältää F1-hybridin naispuoliset sukusolut ja oikealla - miespuoliset sukusolut. Hilan sisäneliöt sisältävät geenien yhdistelmät, jotka syntyvät niiden sulautuessa, ja kutakin genotyyppiä vastaavan fenotyypin. Jos sukusolut sijoitetaan hilaan kaaviossa esitetyssä järjestyksessä, niin hilassa voit havaita järjestyksen genotyyppien järjestelyssä: kaikki homotsygootit sijaitsevat yhtä diagonaalia pitkin ja kahden geenin heterotsygootit (diheterotsygootit) sijaitsevat sitä pitkin. toinen. Kaikki muut solut ovat monoheterotsygoottien (yhden geenin heterotsygoottien) käytössä.
Katkaisu F2:ssa voidaan esittää käyttämällä fenotyyppisiä radikaaleja, so. joka ei osoita koko genotyyppiä, vaan vain geenejä, jotka määräävät fenotyypin. Tämä merkintä näyttää tältä:

Viivat radikaaleissa tarkoittavat, että toiset alleeliset geenit voivat olla joko hallitsevia tai resessiivisiä, ja fenotyyppi on sama.
Dihybridiristeysjärjestelmä
(Punnet-ruudukko)

| AB | Ab | aB | ab | |
| AB | AABB keltainen Ch. |
AABb keltainen Ch. |
AaBB keltainen Ch. |
AaBb keltainen Ch. |
| Ab | AABb keltainen Ch. |
AAbb keltainen rypistyä |
AaBb keltainen Ch. |
Aabb keltainen rypistyä |
| aB | AaBB keltainen Ch. |
AaBb keltainen Ch. |
aaBB vihreä Ch. |
aaBb vihreä Ch. |
| ab | AaBb keltainen Ch. |
Aabb keltainen rypistyä |
aaBb vihreä Ch. |
aabb |
F2-genotyyppien kokonaismäärä Punnettin hilassa on 16, mutta niitä on 9 erilaista, koska jotkut genotyypit toistuvat. Eri genotyyppien esiintymistiheys kuvataan säännöllä:
F2-dihybridiristeyksessä kaikki homotsygootit esiintyvät kerran, monoheterotsygootit kaksi kertaa ja diheterotsygootit neljä kertaa. Punnett-ruudukko sisältää 4 homotsygoottia, 8 monoheterotsygoottia ja 4 diheterotsygoottia.
Segregaatio genotyypin mukaan vastaa seuraavaa kaavaa:
1AABB: 2AABBb: 1AAbb: 2AaBB: 4AaBBb: 2Aabb: 1aaBB: 2aaBBb: 1aabb.
Lyhennettynä 1:2:1:2:4:2:1:2:1.
F 2 -hybrideistä vain kaksi genotyyppiä toistaa vanhempien muotojen genotyypit: AABB Ja aabb; muissa tapauksissa tapahtui vanhempien geenien rekombinaatio. Se johti kahden uuden fenotyyppiluokan syntymiseen: keltaiset ryppyiset siemenet ja vihreät sileät siemenet.
Analysoituaan dihybridiristeytyksen tulokset jokaiselle merkkiparille erikseen, Mendel loi kolmannen mallin: eri merkkiparien periytymisen riippumaton luonne ( Mendelin III laki). Riippumattomuus ilmaistaan siinä, että kunkin ominaisuusparin jakautuminen vastaa monohybridiristeytyskaavaa 3: 1. Näin ollen dihybridiristeytys voidaan esittää kahdeksi samanaikaisesti esiintyväksi monohybridiksi.
Kuten myöhemmin todettiin, riippumaton perinnöllisyystyyppi johtuu geenien lokalisoinnista homologisten kromosomien eri pareihin. Mendelin segregaation sytologinen perusta on kromosomien käyttäytyminen solunjakautumisen aikana ja sitä seuraava sukusolujen fuusio hedelmöityksen aikana. Meioosin pelkistysjakauman profaasissa I homologiset kromosomit konjugoituvat, ja sitten anafaasissa I ne hajoavat eri napoille, minkä vuoksi alleeliset geenit eivät pääse samaan sukusoluun. Kun ne eroavat, ei-homologiset kromosomit yhdistyvät vapaasti keskenään ja siirtyvät napoihin eri yhdistelmissä. Tämä määrää sukusolujen geneettisen heterogeenisyyden ja niiden hedelmöitysprosessin aikana tapahtuvan fuusioitumisen jälkeen tsygoottien geneettisen heterogeenisyyden ja sen seurauksena jälkeläisten genotyyppisen ja fenotyyppisen monimuotoisuuden.
Eri ominaisuusparien itsenäinen periytyminen tekee erotuskaavojen laskemisen helpoksi di- ja polyhybridiristeyksissä, koska ne perustuvat yksinkertaisiin monohybridiristikavoihin. Laskettaessa käytetään todennäköisyyslakia (kahden tai useamman ilmiön esiintymisen todennäköisyys samanaikaisesti on yhtä suuri kuin niiden todennäköisyyksien tulo). Dihybridiristi voidaan hajottaa kahdeksi ja trihybridiristi kolmeksi itsenäiseksi monohybridiristeykseen, joissa kussakin kahden eri ominaisuuden ilmentymisen todennäköisyys F 2:ssa on 3:1. Siksi fenotyypin jakamisen kaava F 2 -dihybridiristi on:
(3: 1) 2 = 9: 3: 3: 1,
kolmihybridi (3:1) 3 = 27:9:9:9:3:3:3:1 jne.
F2-polyhybridiristityksen fenotyyppien lukumäärä on yhtä suuri kuin 2 n, missä n on niiden ominaisuusparien lukumäärä, joissa emoyksilöt eroavat toisistaan.
Kaavat hybridien muiden ominaisuuksien laskemiseksi on esitetty taulukossa 1.
Taulukko 1. Segregaation kvantitatiiviset mallit hybridijälkeläisissä
erilaisiin risteyksiin
| Määrälliset ominaisuudet | Ylityksen tyyppi | ||
| monohybridi | dihybridi | polyhybridi | |
| Hybridin F 1 muodostamien sukusolujen määrä | 2 | 2 2 | 2n |
| Sukusolujen yhdistelmien lukumäärä F 2:n muodostumisen aikana | 4 | 4 2 | 4n |
| Fenotyyppien lukumäärä F 2 | 2 | 2 2 | 2n |
| Genotyyppien lukumäärä F 2 | 3 | 3 2 | 3 |
|
Fenotyypin jakautuminen F 2:ssa |
3: 1 | (3: 1) 2 | (3:1)n |
| Segregaatio genotyypin mukaan F 2:ssa | 1: 2: 1 | (1: 2: 1) 2 | (1:2:1)n |
Mendelin löytämien periytymismallien ilmentyminen on mahdollista vain tietyissä olosuhteissa (kokeilijasta riippumatta). He ovat:
- Yhtä todennäköistä sukusolujen muodostumista kaikkien lajikkeiden hybridoomien kautta.
- Kaikki mahdolliset sukusolujen yhdistelmät hedelmöitysprosessin aikana.
- Kaikkien tsygoottien yhtäläinen elinkelpoisuus.
Jos nämä edellytykset eivät täyty, hybridijälkeläisten erottelun luonne muuttuu.
Ensimmäinen ehto voi rikottua johtuen yhden tai toisen sukusolutyypin elinkelpoisuudesta, mahdollisesti eri syistä, esimerkiksi toisen geenin kielteisestä vaikutuksesta, joka ilmenee gameettitasolla.
Toista ehtoa rikotaan selektiivisen hedelmöityksen tapauksessa, jossa tietyntyyppiset sukusolut fuusioituvat ensisijaisesti. Lisäksi sukusolu, jolla on sama geeni, voi käyttäytyä eri tavalla hedelmöitysprosessin aikana riippuen siitä, onko se naaras vai mies.
Kolmas ehto rikotaan yleensä, jos hallitsevalla geenillä on tappava vaikutus homotsygoottisessa tilassa. Tässä tapauksessa F 2 -monohybridiristeytys hallitsevien homotsygoottien kuoleman seurauksena AA 3:1-jakauman sijasta havaitaan 2:1-jako. Esimerkkejä tällaisista geeneistä ovat: kettujen platinaturkin värin geeni, shirazin lampaiden harmaan turkin värin geeni. (Lisätietoja seuraavassa luennossa.)
Syy poikkeamiseen Mendelin erottelukaavoista voi olla myös piirteen epätäydellinen ilmentymä. Geenien toiminnan ilmentymisaste fenotyypissä ilmaistaan termillä ekspressiivisyys. Joillekin geeneille se on epävakaa ja erittäin riippuvainen ulkoisista olosuhteista. Esimerkkinä on Drosophilan mustan vartalon värin resessiivinen geeni (mutaatioeebenpuu), jonka ilmentyvyys riippuu lämpötilasta, minkä seurauksena tälle geenille heterotsygoottisilla yksilöillä voi olla tumma väri.
Mendelin löytämä perinnönlaki oli yli kolme vuosikymmentä genetiikan kehitystä edellä. Hänen aikalaisensa, mukaan lukien Charles Darwin, eivät ymmärtäneet ja arvostaneet kirjoittajan julkaisemaa teosta "Kokemus kasvihybrideistä". Pääsyy tähän on se, että Mendelin työn julkaisuhetkellä kromosomeja ei ollut vielä löydetty eikä solunjakautumisprosessia, joka, kuten edellä mainittiin, muodosti Mendelin kuvioiden sytologisen perustan, ei ollut vielä kuvattu. Lisäksi Mendel itse epäili löytämiensä kuvioiden yleismaailmallisuutta, kun hän K. Nägelin neuvosta alkoi tarkistaa toisesta esineestä - haukkaruokasta - saatuja tuloksia. Koska Mendel ei tiennyt, että haukkanokka lisääntyy partenogeneettisesti ja siksi siitä on mahdotonta saada hybridejä, Mendel oli täysin masentunut kokeiden tuloksista, jotka eivät mahtuneet hänen lakiensa kehykseen. Epäonnistumisen vaikutuksen alaisena hän luopui tutkimuksestaan.
Tunnustus tuli Mendelille aivan 1900-luvun alussa, kun vuonna 1900 kolme tutkijaa - G. de Vries, K. Correns ja E. Cermak - julkaisi itsenäisesti tutkimustuloksiaan toistaen Mendelin kokeita ja vahvisti hänen tutkimustensa oikeellisuuden. johtopäätökset. Koska tähän mennessä mitoosi, lähes kokonaan meioosi (sen täydellinen kuvaus valmistui vuonna 1905), sekä hedelmöitysprosessi oli kuvattu kokonaan, tutkijat pystyivät yhdistämään Mendelin perinnöllisten tekijöiden käyttäytymisen kromosomien käyttäytymiseen solun aikana. jako. Mendelin lakien uudelleen löytämisestä tuli lähtökohta genetiikan kehitykselle.
1900-luvun ensimmäinen vuosikymmen. siitä tuli mendelismin voittomarssin aika. Mendelin löytämät kuviot vahvistettiin sekä kasvi- että eläinobjektien erilaisten ominaisuuksien tutkimuksessa. Syntyi ajatus Mendelin lakien universaalisuudesta. Samaan aikaan alkoi kertyä tosiasioita, jotka eivät mahtuneet näiden lakien kehykseen. Mutta hybridologinen menetelmä mahdollisti näiden poikkeamien luonteen selvittämisen ja Mendelin johtopäätösten oikeellisuuden vahvistamisen.
Kaikki Mendelin käyttämät hahmoparit periytyivät täydellisen dominanssin tyypin mukaan. Tässä tapauksessa heterotsygootin resessiivisellä geenillä ei ole vaikutusta, ja heterotsygootin fenotyypin määrää yksinomaan hallitseva geeni. Kuitenkin suuri määrä kasveissa ja eläimissä olevia ominaisuuksia periytyy epätäydellisen dominanssin tyypin mukaan. Tässä tapauksessa F 1 -hybridi ei täysin toista yhden tai toisen vanhemman ominaisuutta. Ominaisuuden ilmentyminen on keskitasoa, jossa on suurempi tai pienempi poikkeama suuntaan tai toiseen.
Esimerkki epätäydellisestä dominanssista voi olla kukkien keskitasoinen vaaleanpunainen väri yökauneushybrideissä, jotka saadaan risteyttämällä kasveja hallitsevan punaisen ja resessiivisen valkoisen värin kanssa (katso kaavio).
Kaavio epätäydellisestä dominoinnista kukkavärin perinnössä yön kauneudessa

Kuten kaaviosta voidaan nähdä, ensimmäisen sukupolven hybridien yhtenäisyyslaki pätee risteykseen. Kaikilla hybrideillä on sama väri - vaaleanpunainen - geenin epätäydellisen dominanssin seurauksena A. Toisessa sukupolvessa eri genotyypeillä on sama esiintymistiheys kuin Mendelin kokeessa, ja vain fenotyyppinen erottelukaava muuttuu. Se vastaa genotyypin erottelukaavaa - 1: 2: 1, koska jokaisella genotyypillä on oma ominaisuutensa. Tämä seikka helpottaa analyysiä, koska analyyttistä risteystä ei tarvita.
Heterotsygootissa on toisenlainen alleelisten geenien käyttäytyminen. Sitä kutsutaan kodominanssiksi, ja se kuvataan tutkimuksessa veriryhmien periytymisestä ihmisillä ja useilla kotieläimillä. Tässä tapauksessa hybridillä, jonka genotyyppi sisältää molemmat alleeliset geenit, on molemmat vaihtoehtoiset ominaisuudet yhtä lailla. Kodominanssia havaitaan periessään A-, B-, 0-järjestelmän veriryhmiä ihmisillä. Ihmisissä, joilla on ryhmä AB(IV ryhmä) veressä on kaksi erilaista antigeeniä, joiden synteesiä ohjaa kaksi alleelista geeniä.
1. Kuvaile G. Mendelin ensimmäinen ja toinen laki.
Mendelin ensimmäinen laki (ensimmäisen sukupolven hybridien yhtenäisyyslaki) sanoo, että kun risteytetään alleelisten ominaisuuksien suhteen erilaisia organismeja, vain yksi niistä ilmestyy ensimmäisen sukupolven hybrideihin - hallitseva, kun taas vaihtoehto, resessiivinen, pysyy piilossa.
Mendelin toinen laki (segregaatiolaki) sanoo, että kun kaksi ensimmäisen sukupolven hybridiä risteytetään keskenään, molemmat alkuperäisen vanhempainmuodon ominaisuudet näkyvät tietyssä suhteessa toisessa sukupolvessa.
2. Mitä yhtäläisyyksiä ja eroja G. Mendelin kolmannella lailla ja T. Morganin lailla on?
Samankaltaisuus piilee siinä, että sekä Mendelin kolmas laki että Morganin laki käsittelevät kahden erilaisen ominaisuuden periytymistä. Erot ovat siinä, että Mendelin laissa kyseessä on itsenäinen perinnöllisyys, kun taas Morganin laissa puhutaan ominaisuuksien linkitetystä (yhteisestä) periytymisestä.
3. Mikä on perinnöllisyyden ja vaihtelevuuden rooli elävässä luonnossa?
Perinnöllisyyden ansiosta lajien säilyminen varmistetaan merkittävinä ajanjaksoina (jopa satoja miljoonia vuosia). Ympäristöolosuhteet kuitenkin muuttuvat (joskus merkittävästi) ajan myötä, ja tällaisissa tapauksissa vaihtelevuus, joka johtaa yksilöiden monimuotoisuuteen lajin sisällä, varmistaa sen selviytymisen. Jotkut yksilöt osoittautuvat paremmin sopeutuneiksi uusiin olosuhteisiin, ja tämä antaa heille mahdollisuuden selviytyä. Lisäksi vaihtelevuuden ansiosta lajit voivat laajentaa elinympäristönsä rajoja ja kehittää uusia alueita.
4. Ovatko G. Mendelin lait universaaleja ja soveltuvatko ne ihmisiin?
Joo. Mendelin lait ovat yleismaailmallisia ja koskevat kaikkia eläviä organismeja.
5. Kuvaile käsitteiden "geeni", "alleeli", "risteytys" välistä suhdetta.
Geenit ovat DNA-molekyylin osa, joka vastaa organismin yhden proteiinin tai RNA:n rakentamisesta. Alleelit ovat saman geenin tilan muotoja, jotka miehittävät identtisiä lokuksia homologisissa kromosomeissa ja aiheuttavat fenotyyppisiä eroja samassa ominaisuudessa. Crossing over on homologisten kromosomien homologisten osien risteytys, keskinäinen vaihto.
6. Mikä on mutaatio? Milloin ja missä mutaatioita tapahtuu?
Mutaatiot ovat äkillisiä periytyviä muutoksia geneettisessä materiaalissa, jotka aiheuttavat muutoksia organismin ominaisuuksissa ja ominaisuuksissa. Mutaatiot voivat olla luonnollisia, spontaaneja, eli tapahtuvia tahattomasti tai keinotekoisia, jotka tapahtuvat, kun organismit altistuvat erilaisille tekijöille - mutageeneille.
7. Millaisia ylityksiä G. Mendel tutki?
Monohybridi- ja dihybridiristeys.
8. Mikä on geenien moninkertaisen toiminnan erikoisuus?
Pleiotropia on useiden geenitoimintojen ilmiö. Se ilmenee yhden geenin kyvyssä vaikuttaa useisiin fenotyyppisiin piirteisiin. Esimerkki: geeni, joka vastaa punaisen pigmentin muodostumisesta kukassa, edistää sen ilmestymistä varteen ja lehtiin, aiheuttaa varren pidentymistä ja siementen massan kasvua.
17.06.2010G. Mendelin lait kuvaavat yksittäisten ominaisuuksien periytymisen luonnetta useiden sukupolvien aikana.
Mendelin ensimmäinen laki eli yhtenäisyyden sääntö
Laki johdettiin G. Mendelin keräämien tilastotietojen perusteella risteyttäessään eri hernelajikkeita, joilla oli selviä vaihtoehtoisia eroja seuraavissa ominaisuuksissa:
- siemenen muoto (pyöreä/ei-pyöreä);
- siementen väri (keltainen/vihreä)
- siemenkuori (sileä/ryppyinen) jne.
Ristitettäessä kasveja sileillä ja ryppyisillä siemenillä, kaikki ensimmäisen sukupolven hybridit osoittautuivat sileiksi. Tämä merkki nimettiin hallitseva.
Ristitettäessä homotsygoottisia yksilöitä, jotka eroavat yhden tai useamman vaihtoehtoisen piirteen osalta, kaikki ensimmäisen sukupolven hybridit ovat näissä ominaisuuksissa yhtenäisiä ja samanlaisia kuin emo, jolla on hallitseva ominaisuus.
Jos kyseessä on epätäydellinen dominanssi toisessa sukupolvessa, vain 25 % yksilöistä on fenotyyppisesti samanlaisia kuin hallitsevan piirteen omaava vanhempi. Heterotsygootit ovat fenotyyppisesti erilaisia kuin he. Esimerkiksi punakukkaisten ja valkokukkaisten snapdragon-kasvien jälkeläisistä 25 % on punaisia, 25 % valkoisia ja 50 % vaaleanpunaisia. Testiristeyttämistä käytetään yksilön heterotsygoottisuuden tunnistamiseen tietyn alleelin suhteen. Tätä varten yksilö, jolla on hallitseva piirre (AA? tai Aa?), risteytetään resessiivisen alleelin homotsygoottisen yksilön kanssa. Kun kyseessä on yksilön heterotsygoottisuus, jolla on hallitseva piirre, jälkeläisten erottelu on 1:1:

Mendelin toinen laki tai erottelulaki
Kun ensimmäisen sukupolven heterotsygoottiset hybridit risteytetään keskenään, tämän ominaisuuden pilkkoutuminen havaitaan toisessa sukupolvessa. Tämä jakautuminen on luonnollista tilastollista: 3:1 fenotyypin mukaan ja 1:2:1 genotyypin mukaan.

Riisi. 19. Mypohybridin pilkkomisen sytologinen perusta

Siemenet näkyvät sekä sileillä että ryppyisillä kuorilla.
Mendelin kolmas laki eli itsenäisen periytymisen laki dihybridissä (polyhybridiristeyksessä)
Tämä laki johdettiin tulosten analyysistä, jotka saatiin risteyttämällä yksilöitä, jotka eroavat toisistaan vaihtoehtoisten ominaisuuksien parien suhteen. Esimerkiksi kasvi, joka tuottaa keltaisia, sileitä siemeniä, risteytetään kasvin kanssa, joka tuottaa vihreitä, ryppyisiä siemeniä.

| Sukusolut | AB | Ab | aB | ab |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | ААbb | AaBb | Ааbb |
| aB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
Toisessa sukupolvessa voi esiintyä neljä fenotyyppiä suhteessa 9:3:3:1 ja yhdeksän genotyyppiä.
Analyysin tuloksena havaittiin, että eri alleeliparien geenit ja vastaavat ominaisuudet välittyvät toisistaan riippumatta. Tämä laki on totta:
- diploidisille organismeille;
- geeneille, jotka sijaitsevat eri homologisissa kromosomeissa;
- homologisten kromosomien riippumattomalla erolla meioosissa ja niiden satunnaisessa yhdistelmässä hedelmöityksen aikana.
Nämä olosuhteet ovat sytologinen perusta dihybridiristeytykselle.
Samat mallit pätevät polyhybridiristeyksiin.
Mendelin kokeet totesivat perinnöllisen materiaalin diskreettisyyden (epäjatkuvuuden), mikä johti myöhemmin geenien löytämiseen perinnöllisen tiedon perusmateriaalina kantajina.
Sukusolujen puhtaushypoteesi väittää, että sukusolu sisältää yleensä aina vain yhden tietyn parin homologisista kromosomeista. Segregaatio on seurausta eri alleeleita kantavien sukusolujen satunnaisesta yhdistelmästä.
Koska tapahtumat ovat satunnaisia, kuvio on luonteeltaan tilastollinen, ts. määräytyy suuresta määrästä yhtä todennäköisiä tapahtumia - eri (tai identtisiä) vaihtoehtoisia geenejä kantavien sukusolujen kohtaamisia.
 Verbin aikamuodot eivät muutu henkilökohtaisesti
Verbin aikamuodot eivät muutu henkilökohtaisesti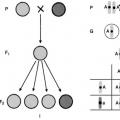 Mendelin lakien täyttymisen ehdot
Mendelin lakien täyttymisen ehdot Jakava pehmeä merkki
Jakava pehmeä merkki